



.png")
1抗HBV药物
1.1 直接抗病毒药物
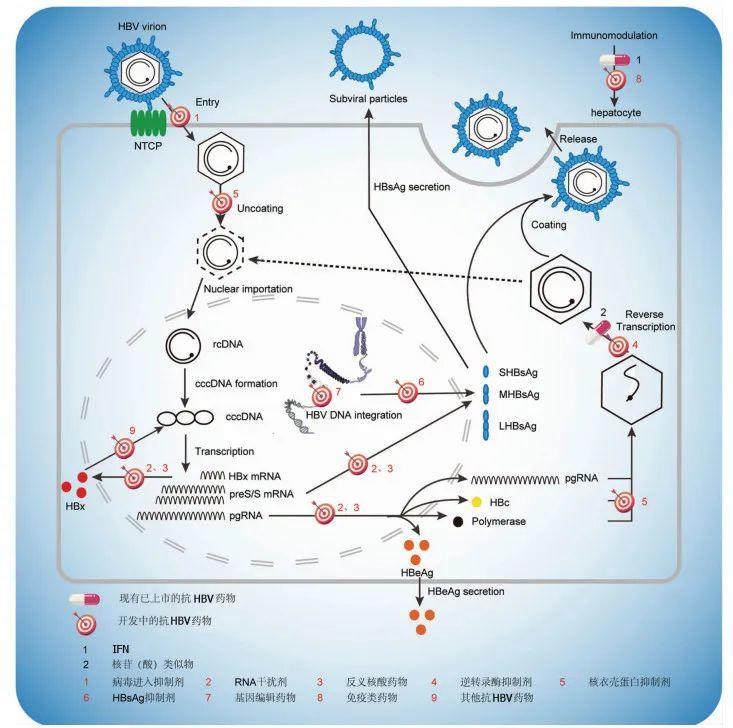
1.1.1 病毒进入抑制剂
1.1.2 RNA干扰剂
1.1.3 反义核酸药物
1.1.4 逆转录酶抑制剂
1.1.5 核衣壳蛋白抑制剂
1.1.6 HBsAg抑制剂
1.1.7 基因编辑药物
1.2 免疫类药物
1.2.1 治疗性疫苗
1.2.2 先天免疫系统激活药物
1.2.3 单克隆抗体
1.2.4 检查点抑制剂
1.2.5 其他免疫途径
1.3 其他抗HBV药物
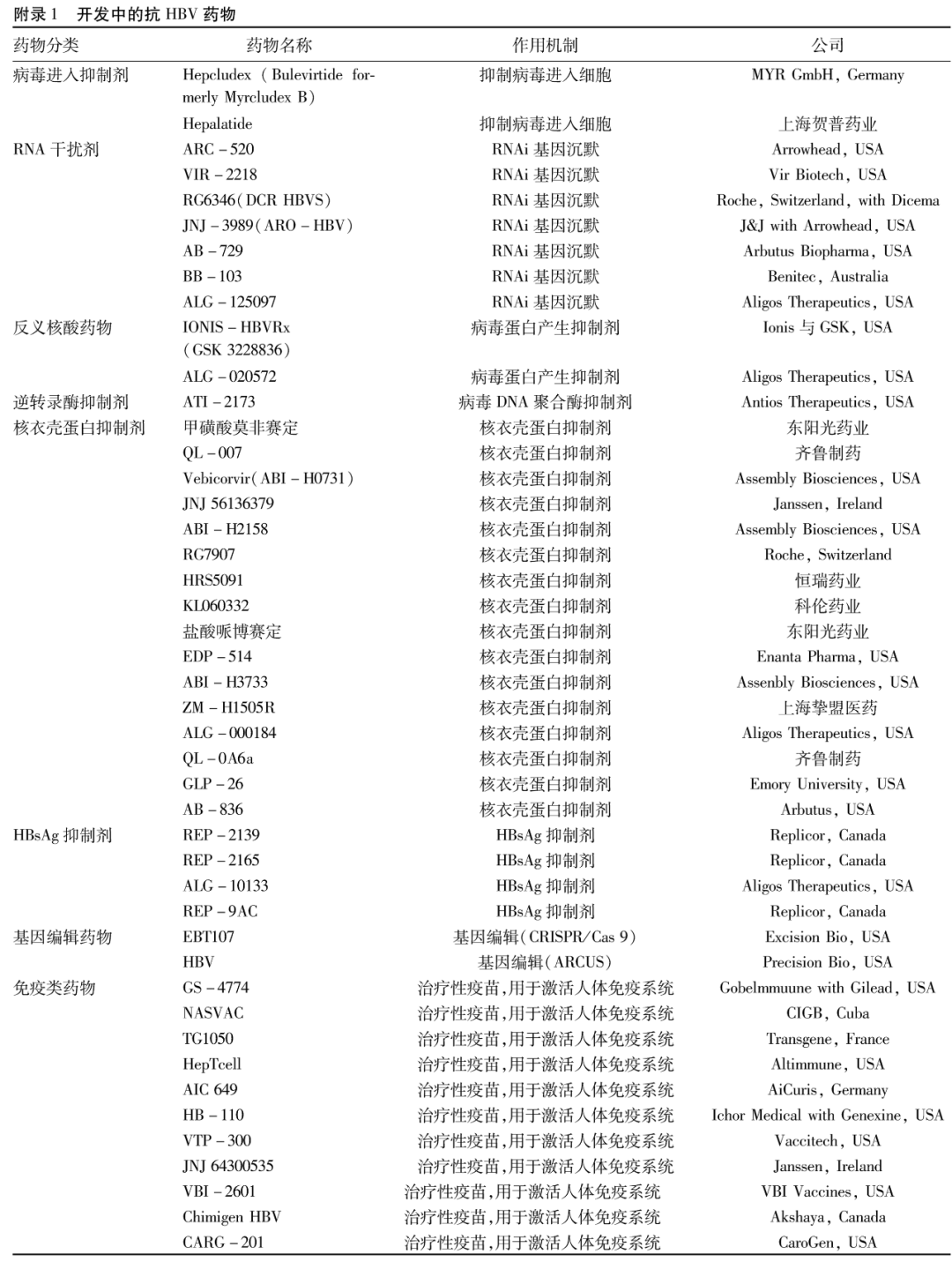
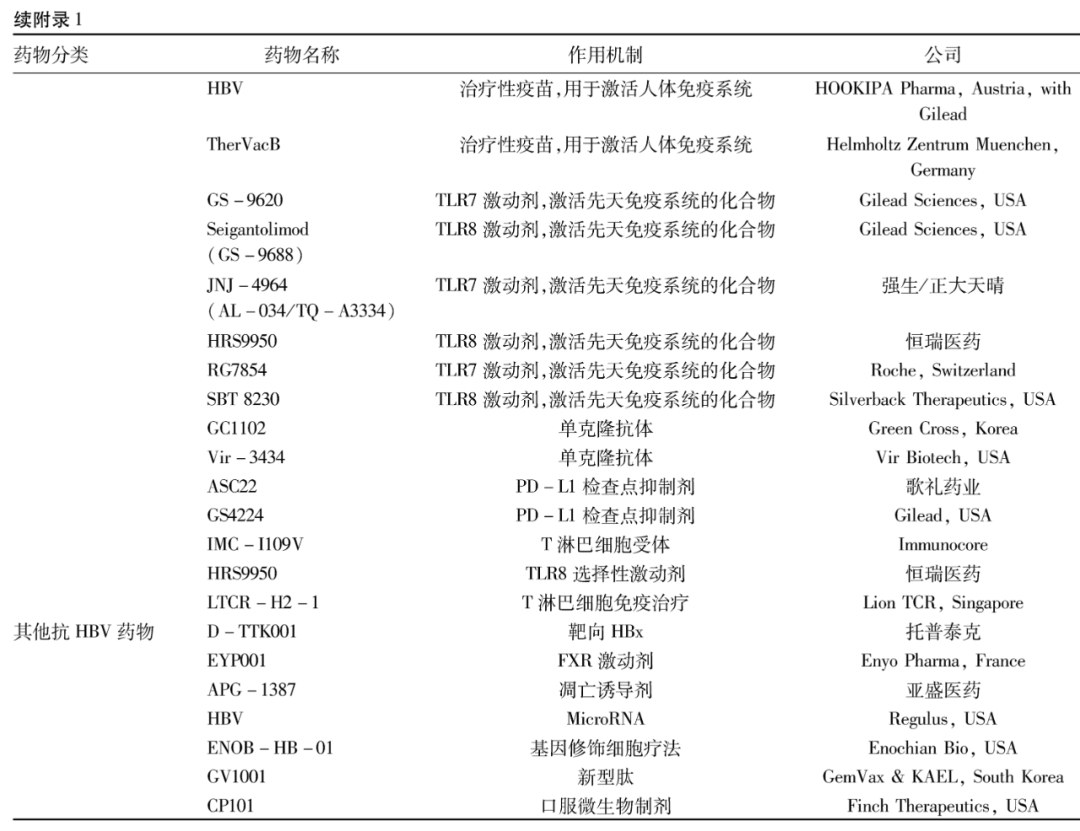
2阻断宿主因子与病毒相互作用的药物
2.1 靶向cccDNA甲基化
2.2 靶向cccDNA的组蛋白修饰
3抗HBV新药展望
参考文献:
[1]SCHWEITZER A, HORN J, MIKOLAJCZYK RT, et al. Estimations of worldwide prevalence of chronic hepatitis B virus infection: A systematic review of data published between 1965 and 2013[J]. Lancet, 2015, 386(10003): 1546-1555. DOI: 10.1016/S0140-6736(15)61412-X.
[2]HOOFNAGLE JH, LAU D. New therapies for chronic hepatitis B[J]. J Viral Hepat, 1997, 4(Suppl 1): 41-50. DOI: 10.1111/j.1365-2893.1997.tb00159.x.
[3]LIANG TJ, BLOCK TM, MCMAHON BJ, et al. Present and future therapies of hepatitis B: From discovery to cure[J]. Hepatology, 2015, 62(6): 1893-1908. DOI: 10.1002/hep.28025.
[4]SUNG JJ, WONG ML, BOWDEN S, et al. Intrahepatic hepatitis B virus covalently closed circular DNA can be a predictor of sustained response to therapy[J]. Gastroenterology, 2005, 128(7): 1890-1897. DOI: 10.1053/j.gastro.2005.03.009.
[5]WERLE-LAPOSTOLLE B, BOWDEN S, LOCARNINI S, et al. Persistence of cccDNA during the natural history of chronic hepatitis B and decline during adefovir dipivoxil therapy[J]. Gastroenterology, 2004, 126(7): 1750-1758. DOI: 10.1053/j.gastro.2004.03.018.
[6]GOVAN L, WU O, XIN Y, et al. Comparative effectiveness of antiviral treatment for hepatitis B: A systematic review and Bayesian network meta-analysis[J]. Eur J Gastroenterol Hepatol, 2015, 27(8): 882-894. DOI: 10.1097/MEG.0000000000000376.
[7]YE X, ZHOU M, HE Y, et al. Efficient inhibition of hepatitis B virus infection by a preS1-binding Peptide[J]. Sci Rep, 2016, 6: 29391. DOI: 10.1038/srep29391.
[8]KANEKO M, WATASHI K, KAMISUKI S, et al. A novel tricyclic polyketide, vanitaracin A, specifically inhibits the entry of hepatitis B and D viruses by targeting sodium taurocholate cotransporting polypeptide[J]. J Virol, 2015, 89(23): 11945-11953. DOI: 10.1128/JVI.01855-15.
[9]DONG Z, EKINS S, POLLI JE. Structure-activity relationship for FDA approved drugs as inhibitors of the human sodium taurocholate cotransporting polypeptide (NTCP)[J]. Mol Pharm, 2013, 10(3): 1008-1019. DOI: 10.1021/mp300453k.
[10]YAN H, LIU Y, SUI J, et al. NTCP opens the door for hepatitis B virus infection[J]. Antiviral Res, 2015, 121: 24-30. DOI: 10.1016/j.antiviral.2015.06.002.
[11]VOLZ T, ALLWEISS L, BEN MBAREK M, et al. The entry inhibitor Myrcludex-B efficiently blocks intrahepatic virus spreading in humanized mice previously infected with hepatitis B virus[J]. J Hepatol, 2013, 58(5): 861-867. DOI: 10.1016/j.jhep.2012.12.008.
[12]BLANK A, MARKERT C, HOHMANN N, et al. First-in-human application of the novel hepatitis B and hepatitis D virus entry inhibitor myrcludex B[J]. J Hepatol, 2016, 65(3): 483-489. DOI: 10.1016/j.jhep.2016.04.013.
[13]CARTHEW RW, SONTHEIMER EJ. Origins and Mechanisms of miRNAs and siRNAs[J]. Cell, 2009, 136(4): 642-655. DOI: 10.1016/j.cell.2009.01.035.
[14]YUEN MF, SCHIEFKE I, YOON JH, et al. RNA interference therapy with ARC-520 results in prolonged hepatitis B surface antigen response in patients with chronic hepatitis B infection[J]. Hepatology, 2020, 72(1): 19-31. DOI: 10.1002/hep.31008.
[15]GISH RG, YUEN MF, CHAN HL, et al. Synthetic RNAi triggers and their use in chronic hepatitis B therapies with curative intent[J]. Antiviral Res, 2015, 121: 97-108. DOI: 10.1016/j.antiviral.2015.06.019.
[16]BUTI M, ESTEBAN R. Drugs in development for hepatitis B[J]. Drugs, 2005, 65(11): 1451-1460. DOI: 10.2165/00003495-200565110-00001.
[17]LIU J, ZHANG S, WANG Q, et al. Seroepidemiology of hepatitis B virus infection in 2 million men aged 21-49 years in rural China: A population-based, cross-sectional study[J]. Lancet Infect Dis, 2016, 16(1): 80-86. DOI: 10.1016/S1473-3099(15)00218-2.
[18]FUNG J, WONG T, CHOK K, et al. Oral nucleos(t)ide analogs alone after liver transplantation in chronic hepatitis B with preexisting rt204 mutation[J]. Transplantation, 2017, 101(10): 2391-2398. DOI: 10.1097/TP.0000000000001883.
[19]ZHOU J, LIU YY, LIAN JS, et al. Efficacy and safety of tenofovir disoproxil treatment for chronic hepatitis B patients with genotypic resistance to other nucleoside analogues: A prospective study[J]. Chin Med J (Engl), 2017, 130(8): 914-919. DOI: 10.4103/0366-6999.204107.
[20]BERKE JM, DEHERTOGH P, VERGAUWEN K, et al. Capsid assembly modulators have a dual mechanism of action in primary human hepatocytes infected with hepatitis B virus[J]. Antimicrob Agents Chemother, 2017, 61(8). DOI: 10.1128/AAC.00560-17.
[21]KLUMPP K, LAM AM, LUKACS C, et al. High-resolution crystal structure of a hepatitis B virus replication inhibitor bound to the viral core protein[J]. Proc Natl Acad Sci U S A, 2015, 112(49): 15196-15201. DOI: 10.1073/pnas.1513803112.
[22]DEWITT MA, MAGIS W, BRAY NL, et al. Selection-free genome editing of the sickle mutation in human adult hematopoietic stem/progenitor cells[J]. Sci Transl Med, 2016, 8(360): 360ra134. DOI: 10.1126/scitranslmed.aaf9336.
[23]RAMANAN V, SHLOMAI A, COX DB, et al. CRISPR/Cas9 cleavage of viral DNA efficiently suppresses hepatitis B virus[J]. Sci Rep, 2015, 5: 10833. DOI: 10.1038/srep10833.
[24]WANG J, CHEN R, ZHANG R, et al. The gRNA-miRNA-gRNA ternary cassette combining CRISPR/Cas9 with RNAi approach strongly inhibits hepatitis B virus replication[J]. Theranostics, 2017, 7(12): 3090-3105. DOI: 10.7150/thno.18114.
[25]LOK AS, PAN CQ, HAN SH, et al. Randomized phase Ⅱ study of GS-4774 as a therapeutic vaccine in virally suppressed patients with chronic hepatitis B[J]. J Hepatol, 2016, 65(3): 509-516. DOI: 10.1016/j.jhep.2016.05.016.
[26]MENNE S, TUMAS DB, LIU KH, et al. Sustained efficacy and seroconversion with the Toll-like receptor 7 agonist GS-9620 in the Woodchuck model of chronic hepatitis B[J]. J Hepatol, 2015, 62(6): 1237-1245. DOI: 10.1016/j.jhep.2014.12.026.
[27]LANFORD RE, GUERRA B, CHAVEZ D, et al. GS-9620, an oral agonist of Toll-like receptor-7, induces prolonged suppression of hepatitis B virus in chronically infected chimpanzees[J]. Gastroenterology, 2013, 144(7): 1508-1517, 1517. e1-10. DOI: 10.1053/j.gastro.2013.02.003.
[28]GANE EJ, LIM YS, GORDON SC, et al. The oral toll-like receptor-7 agonist GS-9620 in patients with chronic hepatitis B virus infection[J]. J Hepatol, 2015, 63(2): 320-328. DOI: 10.1016/j.jhep.2015.02.037.
[29]JANSSEN H, BRUNETTO MR, KIM YJ, et al. Safety, efficacy and pharmacodynamics of vesatolimod (GS-9620) in virally suppressed patients with chronic hepatitis B[J]. J Hepatol, 2018, 68(3): 431-440. DOI: 10.1016/j.jhep.2017.10.027.
[30]SCHINAZI RF, EHTESHAMI M, BASSIT L, et al. Towards HBV curative therapies[J]. Liver Int, 2018, 38(Suppl 1): 102-114. DOI: 10.1111/liv.13656.
[31]SHIN YW, CHO DH, SONG GW, et al. A New ELISA to overcome the pitfalls in quantification of recombinant human monoclonal Anti-HBs, GC1102, by commercial immunoassays[J]. Biol Proced Online, 2018, 20: 18. DOI: 10.1186/s12575-018-0083-8.
[32]CHEN J, WANG XM, WU XJ, et al. Intrahepatic levels of PD-1/PD-L correlate with liver inflammation in chronic hepatitis B[J]. Inflamm Res, 2011, 60(1): 47-53. DOI: 10.1007/s00011-010-0233-1.
[33]GANE E, VERDON DJ, BROOKS AE, et al. Anti-PD-1 blockade with nivolumab with and without therapeutic vaccination for virally suppressed chronic hepatitis B: A pilot study[J]. J Hepatol, 2019, 71(5): 900-907. DOI: 10.1016/j.jhep.2019.06.028.
[34]BOCK CT, SCHRANZ P, SCHRZÖDER CH, et al. Hepatitis B virus genome is organized into nucleosomes in the nucleus of the infected cell[J]. Virus Genes, 1994, 8(3): 215-229. DOI: 10.1007/BF01703079.
[35]BOCK CT, SCHWINN S, LOCARNINI S, et al. Structural organization of the hepatitis B virus minichromosome[J]. J Mol Biol, 2001, 307(1): 183-196. DOI: 10.1006/jmbi.2000.4481.
[36]DAWSON MA. The cancer epigenome: Concepts, challenges, and therapeutic opportunities[J]. Science, 2017, 355(6330): 1147-1152. DOI: 10.1126/science.aam7304.
[37]GHOSH SK, PERRINE SP, WILLIAMS RM, et al. Histone deacetylase inhibitors are potent inducers of gene expression in latent EBV and sensitize lymphoma cells to nucleoside antiviral agents[J]. Blood, 2012, 119(4): 1008-1017. DOI: 10.1182/blood-2011-06-362434.
[38]BARLOW DP. Methylation and imprinting: From host defense to gene regulation?[J]. Science, 1993, 260(5106): 309-310. DOI: 10.1126/science.8469984.
[39]VIVEKANANDAN P, DANIEL HD, KANNANGAI R, et al. Hepatitis B virus replication induces methylation of both host and viral DNA[J]. J Virol, 2010, 84(9): 4321-4329. DOI: 10.1128/JVI.02280-09.
[40]SHIM YH, YOON GS, CHOI HJ, et al. p16 Hypermethylation in the early stage of hepatitis B virus-associated hepatocarcinogenesis[J]. Cancer Lett, 2003, 190(2): 213-219. DOI: 10.1016/s0304-3835(02)00613-4.
[41]ZHENG DL, ZHANG L, CHENG N, et al. Epigenetic modification induced by hepatitis B virus X protein via interaction with de novo DNA methyltransferase DNMT3A[J]. J Hepatol, 2009, 50(2): 377-387. DOI: 10.1016/j.jhep.2008.10.019.
[42]TROPBERGER P, MERCIER A, ROBINSON M, et al. Mapping of histone modifications in episomal HBV cccDNA uncovers an unusual chromatin organization amenable to epigenetic manipulation[J]. Proc Natl Acad Sci U S A, 2015, 112(42): e5715-e5724. DOI: 10.1073/pnas.1518090112.
[43]ZHANG W, CHEN J, WU M, et al. PRMT5 restricts hepatitis B virus replication through epigenetic repression of covalently closed circular DNA transcription and interference with pregenomic RNA encapsidation[J]. Hepatology, 2017, 66(2): 398-415. DOI: 10.1002/hep.29133.




推荐阅读
中国好CMO | 71位首席医学官!庆祝第17个国际临床试验日 骨关节炎药物简析 | 我们将如何老去——带着疼痛的关节还是矫健的身躯? -
国内PARP抑制剂三足鼎立局面将被打破,百济神州帕米帕利获批在即


 点击这里,欣赏更多精彩内容!
点击这里,欣赏更多精彩内容!
本篇文章来源于微信公众号:药时代
发布者:药时代,转载请首先联系contact@drugtimes.cn获得授权
 为好文打赏 支持药时代 共创新未来!
为好文打赏 支持药时代 共创新未来! 