北京时间 3月30日深夜,FDA召开了关于Amylyx 的重磅ALS药物AMX0035 的外部专家咨询委员会会议,这场会议对于如何理解 FDA关于「 药物开发和审评决定的考量 」 有很大的意义,尤其是对于中国从事 罕见病药物研发 的企业, 更有参考价值。
由于直播时间在深夜,有很多“同药”没能完整观看,现应广大朋友们的要求,药时代进行直播回放。
( 完整的会议录像,将在近期发布于药时代公众号 上。关注 药时代 ,获取最新消息)
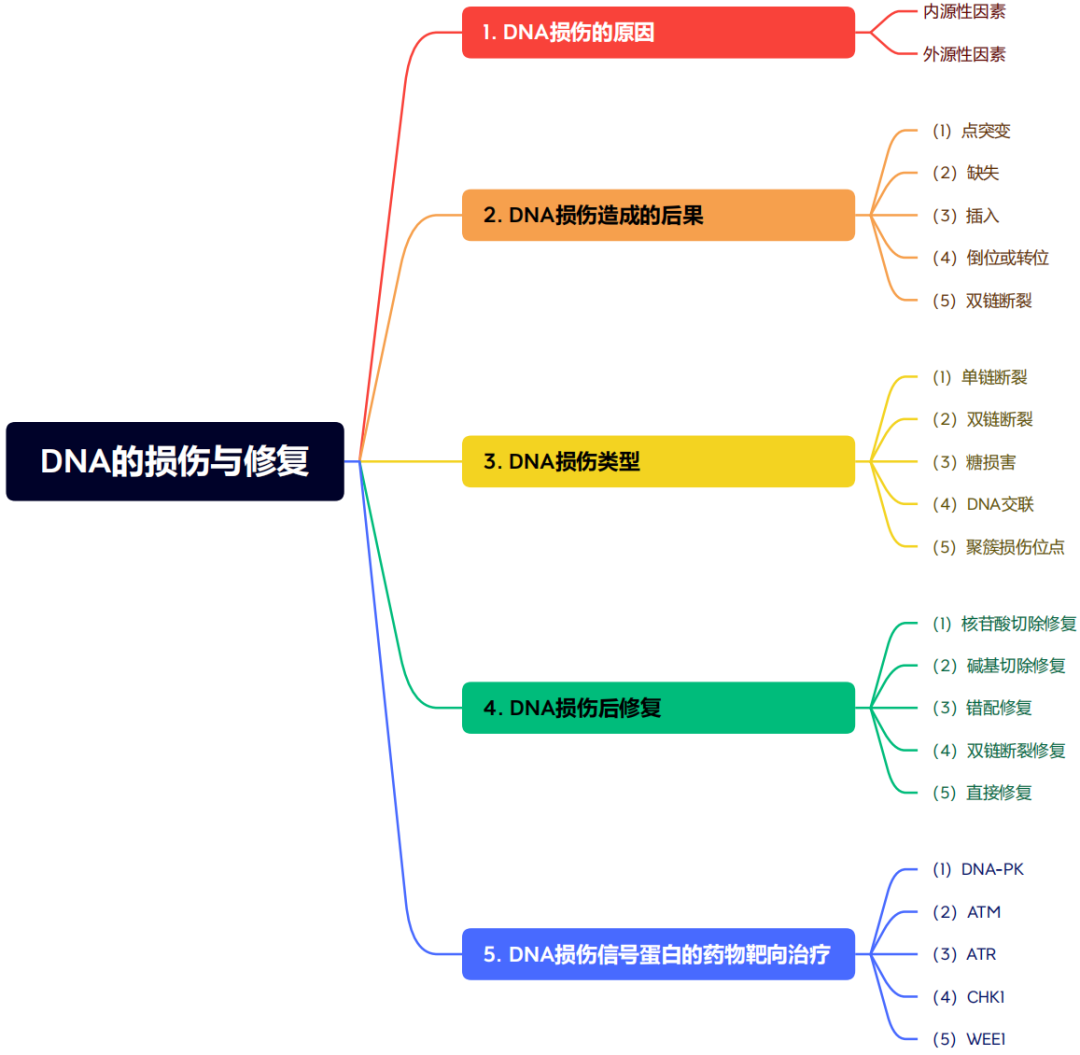
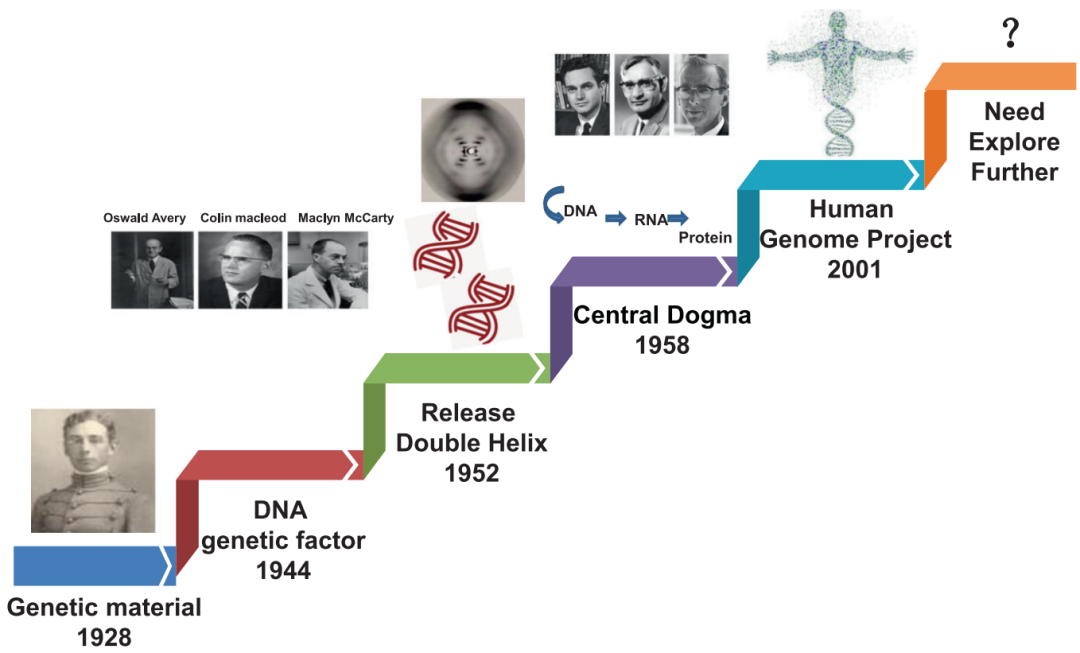
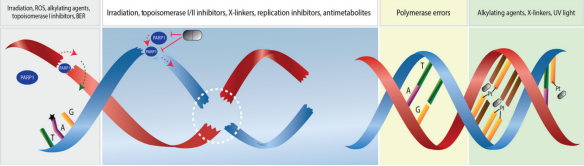
基因组不稳定性是各种癌症的标志,随着DNA损伤的增加积累。放化疗在癌症治疗中的应用通常是基于癌症的这一特性。然而,放疗和化疗也伴随着正常组织损伤等不良反应。靶向癌症治疗具有抑制癌细胞DNA损伤反应的潜力,通过对缺乏特定DNA损伤反应功能的癌症患者进行量身定制治疗。显然,了解DNA损伤修复在癌症中的更广泛作用已成为靶向癌症治疗的基本和有吸引力的策略,特别是在先前科学家发现的基础上提出新的假设或理论,这将对未来有希望的药物新靶点很重要。 DNA损伤修复:DNA损伤修复(repair of DNA damage)是指在多种酶的作用下,生物细胞内的DNA分子受到损伤以后恢复结构的现象。DNA损伤修复的研究有助于了解基因突变的机制,衰老和癌变的原因,还可应用于环境致癌因子的检测。 了解了DNA损伤修复后引入我们今天要介绍的一篇综述,一篇发表2021年7月发表在《Transduction and Targeted Therapy》上的DNA损伤修复应用于癌症靶向治疗的发展历史,机制途径和临床转化的汇总。 1.癌症医学的起源与基于病历分析和流行病学调查的临床发现有关 。被称为流行病学之父的珀西瓦尔·波特观察到阴囊癌在受雇于烟囱清洁工的男孩中发病率很高,并将癌症归因于接触煤烟这是职业暴露于与癌症发展相关的危险因素的第一个证据。 2.在发现DNA结构之前,Theodor Boveri博士在1914年提出了一个著名的理论,即 恶性肿瘤起源于癌细胞 ,癌细胞是通过改变正常细胞而形成的,他通过提出被称为染色体的微小微观体可能在肿瘤细胞中不正常地分布来阐述这一理论。 3.20世纪20年代末,基因突变的主要发现者赫尔曼·穆勒(Hermann Muller)报道说, 将黑腹果蝇暴露在x射线的电离辐射下可能会导致基因的变异,从而导致染色体的畸变 。 4.20世纪30年代,人们观察到,与拥有46条染色体的正常人类细胞相比, 癌细胞中的染色体数量通常变化多端,经常超过46条 。同时,科学家们注意到 癌细胞比正常细胞具有更快、更强的生长能力 。 5.到了20世纪50年代,DNA结构被描述后不久,人们 发现暴露在化学诱变剂(如化学苯)下会导致染色体断裂并增加DNA突变率 。 6. 20世纪80年代 ,对癌变过程进行了描述,描述了由于环境诱变剂的损伤而产生DNA突变的必要条件,以及DNA损伤发生而没有有效修复的情况。因此, DNA损伤反应(DDR)的关键作用被确定 。在接下来的几年里,从DDR过程的研究中发展了许多新的方法,获得了广泛的证据,表明DNA修复,DNA损伤信号和修复、细胞周期检查点、细胞凋亡、复制保真度、DNA重复复制和端粒、均与癌症密切相关基于这些对DNA分子完整性和基因突变过程的研究,DNA突变与细胞癌变之间的联系日益清晰。 7.DNA损伤和致癌的先驱研究者,发现一些烷基化剂,如二氧化丁二烯,可以与 DNA相互作用 ,形成有害的加合物,并最终 破坏DNA作为分子模板的正常作用 ,在过去的几十年里,人们提出并广泛研究了某些癌症基因易受这些因子影响的假说。此后,人们发现化疗药物和放疗可通过诱导DNA损伤有效地治疗各种癌症。在与癌症的战争中,已经开发了许多药剂,也探索了新的技术策略。然而,仍有许多挑战和未解决的问题需要进一步研究,如: (1)肿瘤细胞DNA对化疗药物和放疗反应的详细分子机制; (3)可能的新的和有前途的生物标志物作为新的抑制剂或治疗药物进行研究; (4)最重要的是,DDR的基本生物学机制。有了这些信息,有效的癌症治疗方法可以针对DDR,并最终预防或治愈癌症。 1.DNA分子的自发性损伤(内源性因素) a.DNA的四种碱基各自的异构体间都可以自发的相互变化,这种变化就会使碱基配对间的氢键改变,可使腺嘌呤能配上胞嘧啶、胸腺嘧啶能配上鸟嘌呤等,如果这些配对发生在DNA复制时,就会造成子代DNA序列与亲代DNA不同的错误性损伤腺嘌呤的稀有互变异体与胞嘧啶,或胸腺嘧啶的稀有互变异体与鸟嘌呤的氢链形成导致下一代中C-G配对取代A-T配对。 b.碱基的脱氨基作用,碱基的环外氨基有时会自发脱落,从而胞嘧啶会形成尿嘧啶、腺嘌呤会变成次黄嘌呤、鸟嘌呤会变成黄嘌呤等,遇到复制时,U与A配对、次黄嘌呤与黄嘌呤都与胞嘧啶就会导致自带DNA序列的错误变化。胞嘧啶自发脱氨基的频率约为每个细胞每天190个。 c.脱嘌呤与脱嘧啶:自发的水解可使嘌呤和嘧啶从DNA链的核糖磷酸骨架上脱落下来。 d.碱基修饰和链断裂,细胞呼吸的副产物氧气、过氧化氢等会造成DNA 损伤,能产生胸腺嘧啶乙二醇、羟甲基尿嘧啶等碱基修饰物,还能引起DNA单链断裂等损伤,每个哺乳动物每天DNA单链断裂发生的频率约为5万次。此外体内还可以发生DNA的甲基化,结构的其他变化等,这些损伤的积累可能导致老化。 2.物理因素引起的DNA损伤(外源性因素) : (1)紫外线引起的DNA损伤 ,受到最易吸收的波长(~260nm)的紫外线照射时,主要是使同一条DNA链上相邻的嘧啶以共价键连成二聚体,相邻的两个T、或两个C、或C与T之间都可以环丁基环连成二聚体,其中最容易形成的是TT二聚体。 (2)电离辐射引起的DNA损伤 ,电离辐射损伤DNA有直接和间接的效应,直接效应是DNA 直接吸收射线能量而遭受损伤,间接效应是DNA周围其他分子(只要是水分子)吸收射线能量产生具有很高反应活性的自由基进而损伤DNA。电离辐射可以导致DNA分子的多种变化: a.碱基变化,主要是由OH-自由基引起,包括DNA链上的碱基氧化修饰、过氧化物的形成、碱基环的破坏和脱落等。一般嘧啶比嘌呤更敏感。 b.脱氧核糖上的每个碳原子和羟基上的氢都能与OH-反应,导致脱氧核糖分解,最后引起DNA链断裂。 c.DNA链断裂:DNA双链中一条链断裂称单链断裂;DNA双链在同一处或相近处断裂称为双链断裂。虽然单链断裂发生频率为双链断裂的十到二十倍,但是比较容易修复,对于单倍体细胞(如细菌),一次双链断裂就是致死事件。 3.化学因素引起的DNA损伤(外源性因素) : (1)烷化剂对DNA的损伤 :烷化剂是一类亲电子的化合物,很容易与生物体中大分子的亲核位点起反应。烷化剂的作用可使DNA发生各种类型的损伤: a.碱基烷基化,烷化剂很容易将烷基加到DNA链中嘌呤或嘧啶的N或O上,其中鸟嘌呤的N7和腺嘌呤的N3最容易受攻击,烷基化的嘌呤碱基配对会发生变化,例如鸟嘌呤N7被烷化后就不再与胞嘧啶配对,而改与胸腺嘧啶配对,结果会是G-C变成A-T。 b.碱基脱落,烷化鸟嘌呤的糖苷键不稳定,容易脱落形成DNA上无碱基的位点,复制时可以插入任何核苷酸,造成序列的改变。 c.断链,DNA链的磷酸二脂键上的氧也容易被烷化,结果形成不稳定的磷酸三脂键,容易在糖与磷酸间发生水解,使DNA链断裂。 (2)碱基类似物、修饰剂对DNA的损伤 人工可以合成一些碱基类似物用作促突变剂或抗癌药物 。由于其结构与正常的碱基相似,进入细胞能替代正常的碱基参与到DNA链中而干扰DNA复制合成,导致DNA复制时出现A-T变成G-C。 1.点突变 :DNA上单一碱基的变异。嘌呤替代嘌呤,嘧啶替代嘧啶,成为转换,嘌呤变嘧啶或嘧啶变嘌呤成为颠换。 3.插入 :指一个或一段核苷酸插入到DNA链中。在为蛋白质编码的序列中如缺失及插入的核苷酸数不是3的整倍数,则发生读框移动,使其后所译读的氨基酸序列全部混乱,成为移码突变。 4.倒位或转位 :指DNA链重组使其中一段核苷酸链方向倒置或从一处迁移到另一处。 5.双链断裂 :DNA双链都发生断裂,对于单倍体细胞一个双链断裂就是致死性事件。
四、DNA损伤后修复类型(DNA damage response,DDR)
DNA损伤的后果很严重,所以针对不同类型的损伤。真核生物的DNA修复主要有4种类型:核苷酸切除修复(NER)、碱基切除修复(BER)、错配修复(MMR)和双链断裂修复(DSBR)。
五、DNA损伤引起的细胞反应和DNA损伤修复与癌症的联系
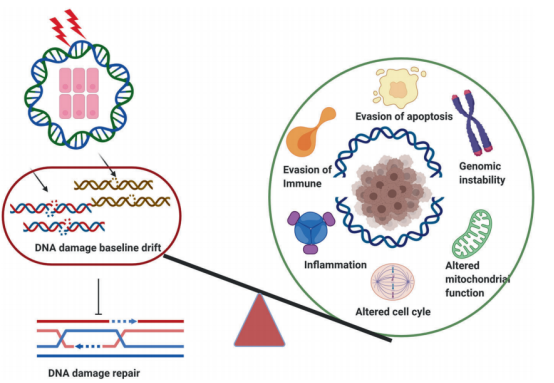
(一)DNA损伤与癌症发生的联系 为了支持生存和繁殖,维持基因组的稳定性是所有细胞的关键优先事项,基因碱基序列的任何异常改变都能破坏细胞的生物学过程,阻碍细胞功能,并可能诱发癌变甚至细胞死亡。具体来说,强有力的证据表明,基因组的不稳定性(原癌基因被连续触发或抗癌基因被抑制)导致了一系列原癌基因的级联反应从而促进癌症发病。
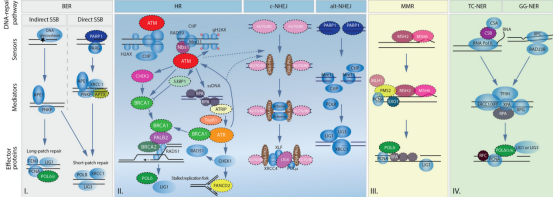
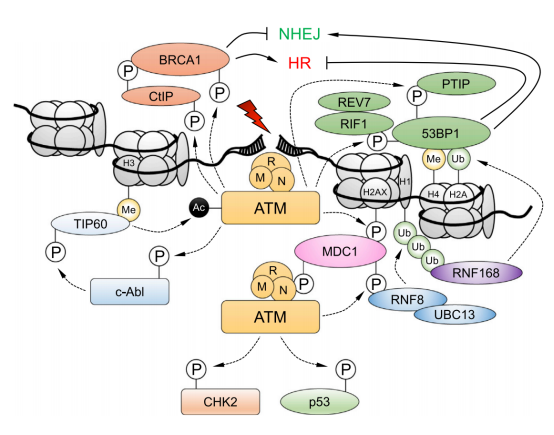
ATM是由DSBs上的MRN复合体招募和激活的。磷酸化-乙酰化级联可通过c-Abl和染色质结合的TIP60维持ATM的激活。ATM磷酸化组蛋白H2AX和MDC1,激活由RNF8和RNF168介导的磷酸化-泛素化信号级联,导致53BP1被招募。ATM使53BP1磷酸化,以促进其效应子的招募,该效应子会被BRCA1和CtIP,以及ATM基板抵消。 在此背景下,EGFR(表皮生长因子受体)、MYC和RAS家族已被普遍认为是原癌基因,而TP53是一种的肿瘤抑制基因。因此,为了减少基因组稳定性受损的遗传失调的可能性, 细胞已经进化出一系列与基因组稳定性相关的信号通路和翻译后修饰来减少这种可能 ,这些通路可以评估DNA代谢的准确性,防止DNA损伤的积累例如,包括ATM (ATM丝氨酸/蛋白激酶)、ATR (ATR丝氨酸/苏氨酸激酶)和DNA-PKcs (DNA依赖蛋白激酶催化亚基)在内的多个家族可以启动哺乳动物细胞中的信号级联。 Monique PCM等人最近综述了泛素化的研究进展,指出 泛素化通过多种酶和蛋白质在调节细胞内稳态中发挥重要作用 。在科学界,这种化合物的复杂功能被称为泛素编码基因组不稳定性,是大多数癌细胞的共同特征。例如,在大多数乳腺癌细胞系中,染色体不稳定性的高比值与有丝分裂纺锤体检查点缺失相关,细胞维持基因组稳定性的分子机制和基因组不稳定性的影响是临床癌症避免相关的基本问题。越来越多的证据表明,DNA双链断裂(DSB)通常是最有害的DNA损伤类型,它损害哺乳动物细胞中基因组的稳定性,许多对抗各种DNA损伤的重要DNA修复功能和过程已经进化。然而, 癌细胞的进化往往与异常的DNA损伤修复功能和过程有关 。例如,在许多癌细胞系中,如套细胞淋巴瘤(MCL), ATM在大约30%到50%的病例中经常发生突变,而这些突变可能与癌症化疗耐药性有关。此外,与细胞周期机制相关的基因在促使癌细胞躲避化疗和放疗的治疗方面发挥了关键作用,大多数杀死癌细胞的措施包括: 尽管有数据显示基因组不稳定性可能与ROS(活性氧自由基)有关,在这篇综述中,作者主要关注DNA损伤修复,因为它是癌症化疗和放疗的主要临床靶点。 (二)癌症中的DNA损伤修复 “DNA损伤基线漂移”假说。与正常细胞相比,癌细胞经历了一个被称为癌变的过程,在这个过程中,DNA损伤导致一系列的基因突变,最终形成大量的细胞,最终生长成肿瘤,称为肿瘤发生 由于DNA双螺旋作为复制和转录模板的作用,它对于保持基因组DNA的稳定性至关重要,如上所述,环境危害和内源性毒素(如自由基)对DNA的损害会损害基因组的稳定性,并引起或促进许多疾病,特别是癌症。 由于DNA分子是基本的遗传物质,确保DNA结构和功能的完整性对支持正常的生命活动和稳定的物种特征至关重要。事实上,当经历内源性或外源性因素的影响时,细胞可以产生各种类型的DNA损伤,包括碱基对改变、DNA复制错误以及DNA双螺旋链的扭曲和断裂常见的外源性因素,特别是某些环境危害,如有毒重金属和电离辐射,发现会造成严重的DNA损伤。 其中内源性物质通常是在体内外源性物质的代谢过程中或在细胞受损和细胞膜完整性丧失后释放出来的。DNA损伤可以通过两种途径发生,即直接作用和间接作用。在直接途径中,内源性或外源性物质直接接触DNA,导致DNA分子化学键断裂,从而改变DNA的结构和活性。 癌症细胞内的内源性应激包括基因转录和复制过程中引起的基因组不稳定性,在间接途径中,内源性或外源性物质激活产物,如自由基,可以破坏DNA。 (三)此前已有几种类型的DNA损伤研究 单链断裂( single – strand break sSSBs)是细胞DNA损伤的主要类型之一,是DNA经氧化应激或者碱基切除修复机制产生的一种常见DNA损伤。体外实验证明,外源H2O2,穿过细胞的膜系统进入核后可直接地损伤 DNA或间接地与二价金属离子(Fe离子和Cu离子)反应生成氧化能力更强的羟自由基(-OH ),进而启动细胞内的一系列代谢事件,使核酶活化、剪切DNA骨架,导致 DNA片段化;H2O2诱导的氧化损伤主要表现为DNA链断裂,并且单链断裂量要比双链断裂量多上千倍。 其中对细胞最有害的损伤和最严重的威胁是双链断裂。没有有效修复或易出错修复的双链断裂可导致致癌或细胞死亡。Lindahl等人报告说,我们的细胞每天可能会遭受大约70000次DNA损伤。这些病变大多数是单链断裂,只有少数是双链断裂,这是较少见的。 大量的研究表明,DNA在氧化应激下表现出大量的碱基和糖损伤,例如鸟嘌呤修饰或7,8-二氢-8-氧-2 -脱氧鸟嘌呤(8-OH-dG)。碱基损伤通常是由氧化应激(如电离辐射诱导的水分子的辐射分解)产生的活性氧间接引起的糖苷损害,如8,5 -环嘌呤-2 -脱氧核苷,可由自由基对糖苷部分的损伤引起。 DNA交联即DNA分子的两条链或同一DNA链的不同区段的侧链间形成的共价相互作用,通常归因于暴露于化学交联剂,如顺铂或产生电离辐射的自由基。在这种类型的损伤中,与DNA修复相关的蛋白质被DNA捕获,导致这些蛋白质粘附在DNA链断裂的5或3端。此外,DNA交联会阻碍一些重要酶的活性,如DNA解旋酶和聚合酶。 聚簇DNA损伤,有时称为DNA多个局部损伤位点,指的是至少有20个碱基对被分开的损伤聚簇DNA损伤通常包括多种损伤,如碱基损伤、碱基部位损伤和单链断裂。然而,与双链断裂不同的是,聚集性DNA损伤的多处损伤可能出现在同一条DNA链上,也可能出现在相反的一条DNA链上,范围很小。总的来说,聚簇性DNA损伤会导致突变频率增加、癌症和细胞死亡。聚簇损伤的机制被描述为一个碱基获得一个电子,然后激活多个电子通路。然而,在癌细胞或正常细胞中聚集DNA损伤是否会产生有益的结果(朋友)或有害的结果(敌人),还需要进一步研究。 (四)尚待解决的DNA损伤发生问题 由于细胞面临着电离辐射或ROS等各种外源性和内源性应激引起的大量DNA损伤,DNA损伤是如何发生的需要更深入的研究。在未来的研究中,仍有许多科学问题有待解决,如: (1)排除了目前已知的DNA损伤类型,可能存在其他尚未发现的新型DNA损伤类型; (2)评估和测量DNA损伤类型和程度的方法,或DNA损伤可视化技术; (3)监测DNA损伤过程,并识别有效的生物标记物,以便及早发现DNA损伤; (4)获得驱动DNA损伤的外源性和内源性应激源的参考值。研究这些问题可能有助于标准化由各种应激引起的DNA损伤。重要的是,在探索这些有趣的问题的同时,将发展出创新的技术和独特的理论模型。 发生癌症时,DNA修复和DNA损伤应答(DDR)通路经常遭到破坏,这是癌症的标志之一。关键DNA修复/DDR基因的种系导致对癌症的易感性和体细胞突变导致癌症中出现较高的突变负担。 合成致死是一个我们在之前的文章中提到的基因组概念,即通常在同一通路中同时破坏两个基因,导致细胞死亡。重要的是,合成致死已被证明发生在基因与药物之间,这一方法已成功地应用于携带DNA修复/DDR缺陷的肿瘤。批准将聚(ADP-核糖)聚合酶(PARP)抑制剂用于治疗BRCA1/2突变的卵巢癌就是例证。在这种情况下,一个基因因突变而失活,另一个因药物而失活。 (二)DNA修复过程 细胞不断地应对因内源性过程(如DNA复制压力)或外源性暴露(如电离辐射和化疗药物)造成的DNA损伤。DNA损伤剂会导致不同类型的DNA损伤,而无法修复损伤会对细胞造成一系列可能的毁灭性后果,包括基因不稳定性和促进肿瘤形成的突变积累。因此,细胞形成了复杂的修复机制,来处理可能出现的不同类型DNA损伤;所有这些都是为了保护基因组的稳定性。 直接修复是最简单的DNA修复形式,因为它主要依赖于单个蛋白的活性,且无需进行核苷酸去除、重新合成或结扎。例如,O6-甲基鸟嘌呤(O6-mG)是一种由烷基化诱变剂引起的有害基因损伤。鸟嘌呤O6位点存在烷基导致G:C转变至A:T和基因转录或DNA复制阻滞。在直接修复过程中,烷基被O6-甲基鸟嘌呤DNA甲基转移酶(MGMT)去除,适当的核苷酸被恢复。 BER负责修复对基因组保真度和稳定性具有重大威胁的较小但高度诱变DNA损伤。这些DNA损伤可由电离辐射以及氧化、甲基化、脱氨等代谢活动或DNA碱基对的自发丢失所产生的内源性诱变引起。BER通路由11种 受损碱基的DNA糖苷酶之一启动。碱基切除后,一组不同的蛋白通过单个碱基(短补丁通路)或多个碱基(长补丁通路)填充暴露的间隙。 3.核苷酸切除修复(NER) 就已经识别和修复损伤的多样性而言,NER可能是最灵活的机制。在这些损伤中,最明显的是UV光诱导的嘧啶二聚体(环丁烷嘧啶二聚体和6-4光产物),其他NER底物包括大体积化学加合物、DNA链间交联和某些形式的氧化损伤。这些类型的损伤会导致DNA双链螺旋扭曲和DNA化学改变,这两者都是NER识别的标志性特征。 NER是一个复杂的多步骤过程,涉及一个由30多种蛋白组成的大网络。首先识别受损碱基,然后解开DNA双链,并通过切除修复复合物去除受损碱基,然后进行填充和结扎。 4.DNA错配修复(MMR) MMR通路在修正复制错误中起着至关重要的作用,例如分别由核苷酸的DNA聚合酶错误掺入和模板滑移导致的碱基-碱基错配和插入或缺失(IDL)。MMR还纠正了由5-甲基胞嘧啶的自发脱氨基作用和基因重组后形成的异源双链核酸分子所产生的错配对或“错配”。 该通路的缺陷导致一种“突变体”细胞表现型,其特征是自发突变频率升高和微卫星不稳定性(MSI)增加。几种人类MMR基因的突变导致对遗传性非息肉病性结直肠癌(HNPCC)的易感性,以及多种表现MSI的散发性肿瘤。 5.双链断裂(DSB)修复
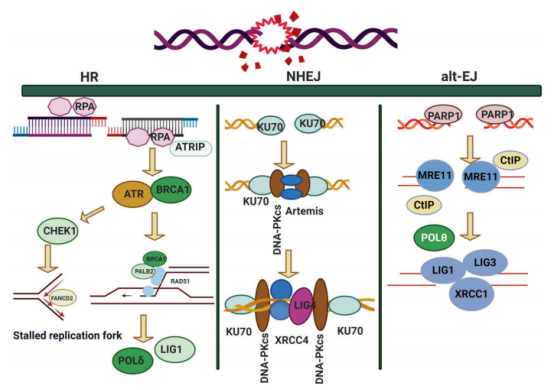
DSB是一种高毒性的基因损伤,对细胞自动调节构成严重威胁,因为它们可以影响转录、复制和染色体分离。DSB由多种外源性因素引起,如电离辐射和某些基因毒性化学物质,以及内源性因素,如活性氧簇、单链DNA断裂的复制和染色体上的机械应力。DSB不同于大多数其他类型的DNA损伤,主要是因为它们影响DNA的双链,因此阻止使用互补链作为模板进行修复(即BER、NER和MMR)。无法修复DSB可导致毁灭性的染色体不稳定性,从而导致基因表达异常和致癌风险增加。 细胞已经进化出两种不同的DSB修复通路:同源重组(HR)和非同源性末端接合(NHEJ): 虽然细胞可能选择使用其中的任何一种通路来修复DSB,但细胞选择其中一种通路而非另一种通路的原因仍然不明。在损伤发生时,选择似乎受到细胞周期阶段的影响。 HR通过修复DSB、间隙和重启停滞的复制交叉来维持基因组的稳定性。这是一种相对缓慢但无错误的通路,它依赖于基因组中存在的同源序列作为模板来替换受损的DNA片段。 近年来,同源重组缺陷(Homologous recombination deficiency,HRD)在癌症生物学中受到了广泛关注,因为在不同癌种中观察到这种 DNA 修复中的缺陷可能对治疗效果产生影响。 DNA发生损伤时,若无法正常通过同源重组(HR)修复通路来修复,即为同源重组缺陷(Homologous recombination deficiency ,HRD)。一些已知的编码同源重组蛋白的基因有:BRCA1、BRCA2、ATM、ATR、BARD1、BLM、RAD51 等。其中, 编码同源重组蛋白的BRAC1 和 BRCA2 的体细胞突变因为涉及遗传性乳腺癌和卵巢癌,成为 HRD中的热门研究对象。 HRD在不同肿瘤中具有普遍性。据研究,HRD存在于13%的肿瘤中,最常见的突变谱系包括卵巢(14.1%),膀胱(9.7%),乳腺(8.0%),子宫内膜(7.4%),前列腺(7.1%)和胰腺(6.5%)。在临床试验中观察到,相比没有HRD的肿瘤,有HRD的肿瘤对铂类以及PARP抑制剂有较好的响应。 与HR不同,NHEJ不需要使用DNA模板(姐妹染色单体)进行修复。相反,NHEJ通过使用不同的核酸酶修饰断裂任意一侧的DNA自由端,使其变得具有兼容性(即3′-羟基和5′-磷酸盐),然后与DNA连接酶连接。与HR相比,NHEJ是一个相对快速但本质上容易出错的过程,过度使用可导致基因重排、缺失和突变,所有这些均可导致复制后细胞更易受到DSB的伤害。 DNA损伤感受器蛋白和DNA损伤信号蛋白可作为一系列抗癌药物的靶点。 (一)DDR感受器蛋白 DDR对修复通路的激活和细胞存活至关重要,而对多种DNA损伤做出应答的DDR感受器蛋白是启动修复的关键。 对于DSB,Ku和MRN是主要感受器蛋白复合物。Ku是一种由Ku70/Ku80构成的蛋白异质二聚体,是NHEJ机制的一部分,能立即与DNA双链断裂结合。在识别和结合DSB后,Ku吸收其他蛋白,以协助进行经典NHEJ修复。 MRN复合物对DSB的初始检测也很重要。在与受损位点结合后,MRN吸收DNA损伤信号激酶ATM,激活并触发一系列信号事件,启动DNA末端切除并促进HR修复。已鉴别出多种其他DNA损伤感受器,包括范可尼贫血核心复合物、错配修复蛋白和核苷酸切除修复蛋白,它们是DNA链间交联、碱基-碱基错配或插入-缺失以及UV诱导光损伤的感受器。 (二)合成致死与DNA损伤修复 PARP-1是一种关键DNA损伤感受器蛋白。由于PARP与BRCA之间存在合成致死关系,已在很大程度上将它作为药物靶点。当同时丧失两个基因的功能(通常在同一通路中)导致细胞死亡时,就会出现合成致死。通过小分子抑制剂(如奥拉帕尼、尼拉帕利、尼拉帕利和他拉唑帕尼)在BRCA突变卵巢癌中成功实现临床PARP靶向治疗,提供了利用合成致死作为治疗策略的原理验证,并已对以DNA损伤应答为靶点的其他疗法进行开发。 (三)DNA损伤信号蛋白的药物靶向治疗 DDR信号蛋白触发各种使DNA损伤信号放大和多样化的翻译后修饰和蛋白复合物装配,以便能够启动适当的应答,并且能够包括:转录变化、细胞周期检查点激活、选择性剪接、参与DNA修复过程,或在大规模损伤的背景下,激活细胞衰老和凋亡通路。 下文将讨论协调DDR信号事件与以此通路为靶点的新药物的主要蛋白 。 NHEJ正常修复DSB需要DNA蛋白激酶(DNA-pk),是贯穿细胞周期各个阶段人类细胞中DSB的主要DNA修复通路。DNA-PK激酶活性的小分子抑制剂使DNA-PK在DNA末端稳定,随后损害NHEJ,还可能干扰其他修复过程,包括通过阻碍DNA末端切除实现的HR。DNA-PK活性的丧失导致细胞增殖减少和胱天蛋白酶介导的细胞死亡启动。 由于与其他激酶的结构相似,与靶向DNA-PK相关的挑战之一是选择性。鉴于它在NHEJ中的作用,经发现在与诱导不依赖复制DSB的药物,如电离辐射和拓扑异构酶2抑制剂(如阿霉素和依托泊苷),联合使用时,以DNA-PK为靶点的药物更为有效。一些小分子目前正处于不同的临床开发阶段,包括MSC2490484A、VX-984和CC-115。 ATM是一种在整个细胞周期中促进DSB修复和响应DSB蛋白激酶。ATM主要通过与MRN复合物的NBS1相互作用激活。它是负责组蛋白H2AX磷酸化的主要激酶,在DSB后迅速发生,并作为DNA修复机制装配的基础。经证明,ATM抑制使细胞对电离辐射和DSB药物(如依托泊苷、喜树碱和阿霉素)非常敏感。 目前,正在评价ATM抑制剂AZD0156的一期试验,作为单药治疗和与PARP抑制剂奥拉帕尼以及其他细胞毒类药物(包括伊立替康)联合使用。 在HR的早期阶段,ATR被ssDNA位点上结合的复制蛋白A(RPA),如停滞的复制叉,或随后在其中一条DNA链发生的5′-3’降解(即DNA末端切除)激活。 Berzosertib(也被称为M6620和VX-970)是首创ATR抑制剂,临床前数据显示肺癌细胞主要对化疗药物敏感。这导致了复制叉崩溃,如顺铂和吉西他滨(体外),并在与顺铂(体内)联合时提高了抗肿瘤活性。 CHK1是ATR下游的蛋白激酶,是S和G2-M细胞周期检查点的关键调节因子。考虑到CHK1在DNA损伤后介导细胞周期阻滞中的作用,当与在DNA复制过程中诱导DNA损伤的药物联合使用时,CHK1抑制剂似乎最为有效,也就不足为奇了。因此,它们的临床开发集中在与此类药物的联合使用上。 另外,MK8776是一种有效的选择性CHK1抑制剂。经证明,单药治疗和与吉西他滨联合使用均具有良好的耐受性。最近,在临床前和临床试验中,CHK1抑制剂prexasertib也显示出具有单药和联合活性。 与CHK1平行作用的WEE1蛋白激酶,通过调节细胞周期蛋白依赖性激酶在激活G2-M检查点中发挥重要作用。但是,与CHK1不同的是,WEE1不直接受DNA损伤的调节,但它是生理细胞周期进程所必需的蛋白激酶。 对于WEE1抑制剂,我们认为其作用机制是通过阻止作为不适当CDK1/CCNB1激活结果的G2-M检查点的激活,从而导致有丝分裂灾变。但是,近期的数据表明,由于CDK2抑制导致DNA复制异常,WEE1抑制也会导致细胞中出现复制依赖性DNA损伤。经证明,首创WEE1激酶抑制剂AZD1775可增加一系列DNA损伤剂的细胞毒性作用,并已在临床前模型中证明了单药活性。最近,一项一期研究发现,AZD1775与拓扑异构酶抑制剂联合使用具有可耐受性,目前已转入二期研究。 虽然NHEJ和HR是主要的DSB修复通路,但选择性同源重组修复机制的重要性正日益得到公认。例如,已知HR缺乏细胞依赖容易出错的微同源介导末端连接(MMEJ)来进行DNA修复和存活。在MMEJ过程中,间隙填充需要POLQ的DNA聚合酶活性(DNA聚合酶θ),且POLQ还通过限制RAD51在切除的DNA末端的积累来防止过度重组。因此,POLQ是一个具有吸引力的药物靶点,尤其在HR缺乏肿瘤中。 正在开发破坏RAD51重组酶家族蛋白-蛋白相互作用的小分子抑制剂,可能对HR缺乏肿瘤特别有效。在近期的一项分析中,确定了14种正处于临床开发中且以DDR为靶点的化合物(除PARP1/2抑制剂以外),并在临床前环境中评价了其他药物。
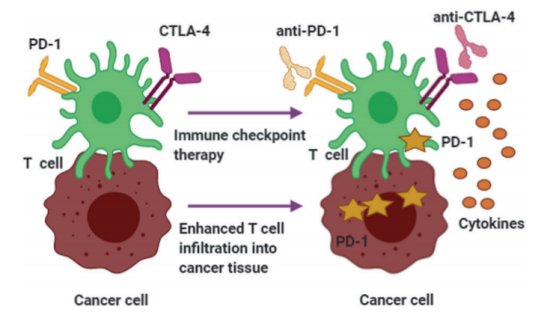
在许多类型的癌症中,有效应对DNA损伤的能力往往会丧失,以这一通路为靶点的药物已在患者亚群中得到了临床验证,如使用PARP抑制剂治疗BRCA1/BRCA2突变的患者。为了提高治疗效果,DDR抑制剂可与以其他DDR蛋白或完全不同的信号通路为靶点的药物联合使用,目的是阻断癌细胞赖以生存的多种通路。 (一)DDR抑制剂与标准治疗药物联合使用 例如,目前临床试验正在检测PARP抑制剂与以DRR通路的其他成员为靶点的小分子的联合使用,包括AZD6738 (ATR)、AZD0156 (ATM)或AZD1775 (WEE1),以及ATM缺乏肿瘤中的ATR抑制、细胞周期蛋白E或MYC扩增肿瘤中的靶向WEE1和HRD或NHEJ缺乏肿瘤中的POLQ抑制剂。临床前研究还表明,以DDR蛋白为靶点对治疗癌基因表达失调的肿瘤可能有利,因为强致癌信号可诱导复制压力。 DDR抑制剂也可与标准治疗药物联合使用,如使用PARP抑制剂增强铂类药物的效果,以及评价其他DDR抑制剂(包括CHK1/2和WEE1抑制剂)与放化疗联合使用的其他研究。 (二)DDR抑制剂与免疫检查点抑制剂联合使用 最后,存在重要的科学依据和临床证据表明,DDR和免疫应答相互关联,且具有潜在协同性。随着我们对DNA损伤、DDR与免疫应答之间相互作用理解的不断加深,将其与DDR抑制剂和/或辐射(作为增敏剂)联合使用可能会提高免疫疗法的临床疗效。 例如,当基因损伤未修复时,会导致肿瘤细胞表面突变负荷和新抗原表达的巨大改变。临床前研究表明,在患BRCA突变型卵巢癌的小鼠模型中,联合使用DNA修复靶向治疗与免疫检查点抑制剂可产生协同效应,如双重CTLA-4和PARP阻断,从而降低肿瘤负荷并提高生存率。 在临床环境中,早期研究支持联合使用PARP抑制剂与抗PD/PD-L1药物的安全性。目前正在进行多项试验,探索与其他DDR抑制剂的联合使用,包括以ATR为靶点的DDR抑制剂。 虽然癌症细胞可能受益于DDR通路中的缺陷,但存在其他起作用的DNA修复系统可提高存活率。以DDR为靶点的药物已在患者亚群中得到了临床验证,且正在积极研究不同的组合策略,以抑制癌细胞生存所依赖的多个通路。通过阻碍DNA修复,DDR抑制剂是适用于联合治疗的理想药物,可以提高辐射、化疗和免疫治疗的疗效。 参考文献: [1] Huang, R., Zhou, PK. DNA damage repair: historical perspectives, mechanistic pathways and clinical translation for targeted cancer therapy. Sig Transduct Target Ther 6, 254 (2021). https://doi.org/10.1038/s41392-021-00648-7
[2] Perkhofer L, Gout J, Roger E, Kude de Almeida F, Baptista Simões C, Wiesmüller L, Seufferlein T, Kleger A. DNA damage repair as a target in pancreatic cancer: state-of-the-art and future perspectives. Gut. 2021 Mar;70(3):606-617. doi: 10.1136/gutjnl-2019-319984
发布者:药时代,转载请首先联系contact@drugtimes.cn获得授权








 为好文打赏 支持药时代 共创新未来!
为好文打赏 支持药时代 共创新未来! 