



细胞的质膜是高效的生物屏障,将细胞内容物拒之门内,而将异物拒之门外。尽管分子量<500 且兼具亲水和疏水基团的有机小分子可以通过被动扩散穿过细胞膜,但生物大分子(例如肽、蛋白质和核酸)通常不能穿膜。然而,一些生物分子能够自主进入哺乳动物细胞的细胞质,例如细胞穿透肽(CPP)、非肽细胞穿透分子 (CPM)、细菌蛋白毒素、某些哺乳动物蛋白和病毒和许多合成药物递送载体已被证明以一种或多种内吞机制进入细胞(图1)。
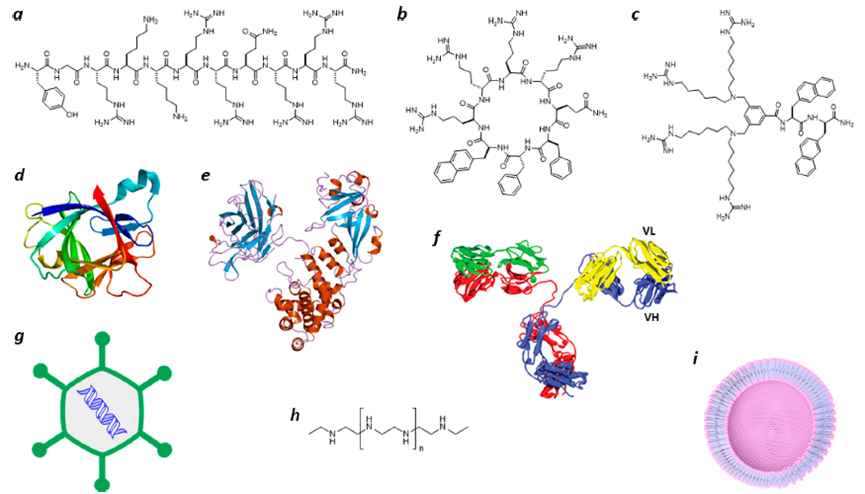
图1 自主跨越细胞质膜或内体膜实现转运的分子或药物
其最初位于膜内体,为了到达胞质溶胶,其中一些分子易位穿过内体膜,这一过程称为“内体逃逸”。其他分子则是直接易位穿过质膜或进一步进入内质网 (ER) 并通过逆行穿过ER膜进入胞质溶胶。折叠的蛋白质也以相反的方向跨过质膜,即从胞质溶胶到细胞外去。在所有这些情况下,生物分子必须穿脂质双层。而生物分子如何在脂质双层中自主易位一直以来是一个谜团,不明确的机制极大地阻碍了穿膜类生物制剂疗法的发展。
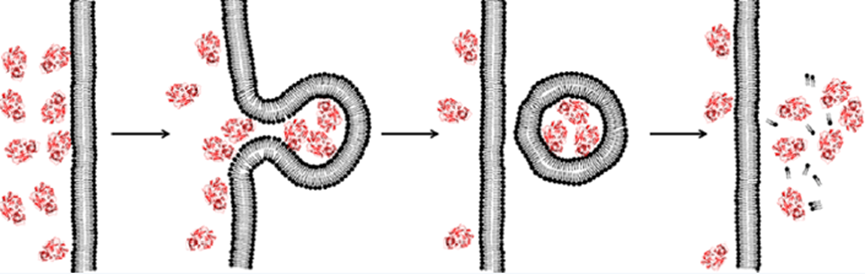
这篇文章首次介绍了一种膜易位机制(囊泡出芽和塌陷(VBC)机制)。通过VBC,不同大小和物理化学性质的生物分子在拓扑上(即从膜的一侧到另一侧)而不是物理上(即不穿过膜)自主地跨细胞膜易位,以期解释之前研究发现的神秘结果。


已经有几种内体逃逸机制被提出,包括质子海绵效应,膜融合,气孔形成,局部膜破坏,和VBC。其中,VBC机制是唯一经过实验验证的机制,而其他假设通常只用于特定模式。图2为环状CPP内吞摄取和内体逃逸的机制。环状CPP直接与质膜磷脂(以及可能的其他膜成分)结合,并通过内吞作用进入早期内体。随着早期内体成熟及内体酸化变为晚期内体,CPP与内体膜结合。这是因为CPP的精氨酸(Arg)侧链在内体酸化期间并不会经历进一步的质子化(pH 6.5- 4.5),质子化可能会发生在磷脂(例如磷酸盐)上。
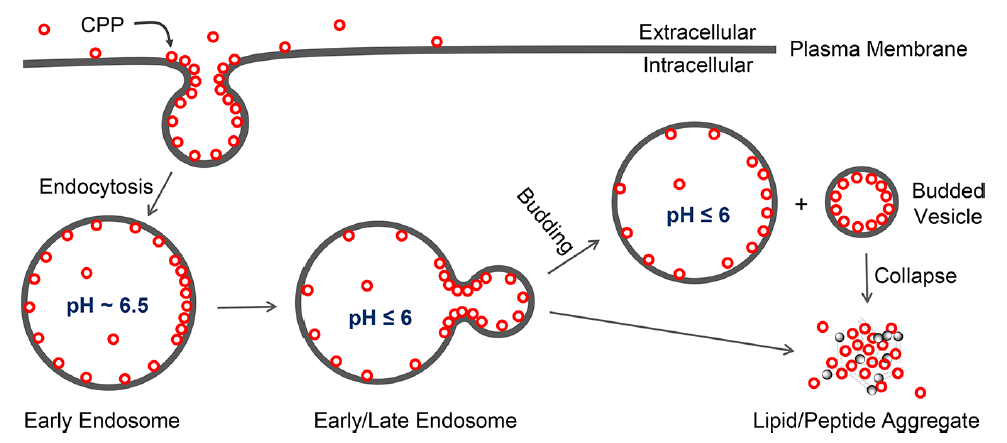
图2 VBC机制对环CPPs的内吞摄取和内体逃逸
Arg的胍基团能够同时与两个相邻的磷酸盐形成氢键(图 3)。基于此,再加上CPP中存在的多个Arg残基(例如,CPP12 中的四个),使CPP能够将磷脂交联成富含CPP的脂质结构域。磷酸盐部分质子化减少静电排斥进而促进磷脂聚集。脂质结构域的形成会在结构域与其周围膜之间产生线张力,从而驱动脂质结构域以囊泡的形式出芽。在囊泡出芽过程中,出芽颈(过渡态)具有负高斯曲率(即在正交方向上同时存在正曲率和负曲率)并且在萌芽之前或之后具有高势能(图 3b)。
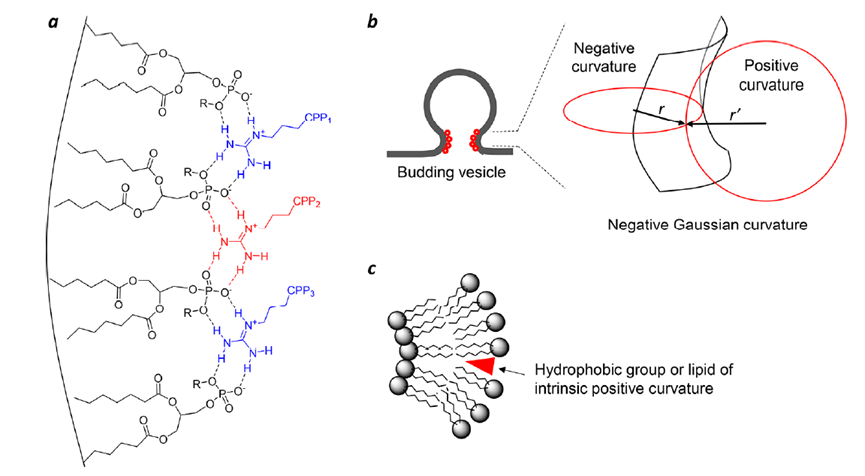
图3. 生物分子对膜结构的影响。
VBC机制的一个独特特征是生物分子在拓扑上穿过膜,但没有物理穿过膜,且内体在每次内体逃逸事件之前、期间和之后都保持完整。相比之下,其他的膜转运机制都涉及细胞膜的破损。
细胞穿透肽
人们普遍认为,在低浓度下,CPPs 主要通过一种或多种能量依赖性内吞机制进入细胞,然后进行内体逃逸。然而在高浓度下,一些 CPPs 也可以不依赖能量直接穿过质膜。
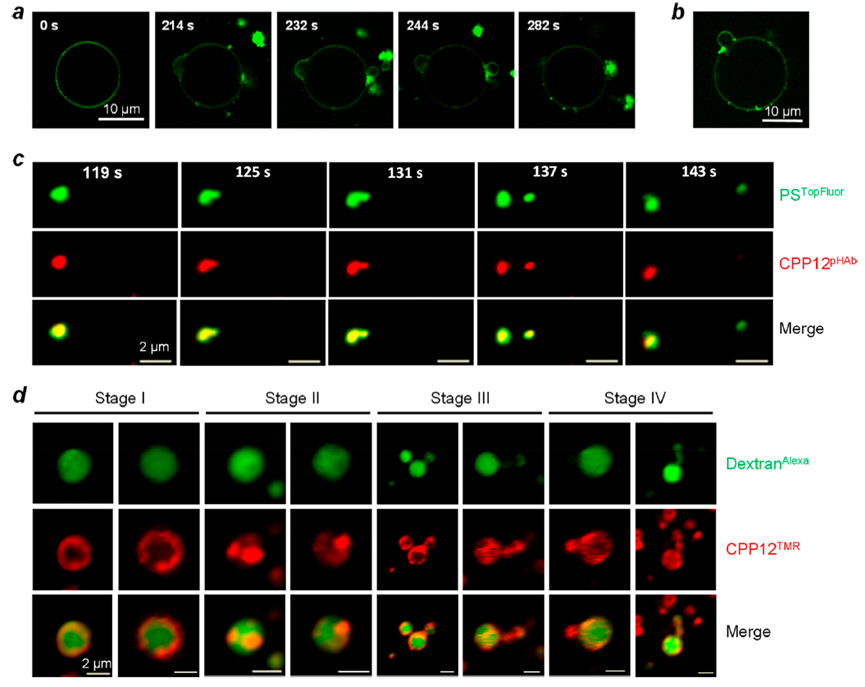
图4.CPP12诱导囊泡出芽并在体外和活细胞中塌陷
这一研究有助于解释VBC机制,(1) CPPs能够输送不同大小(从小分子到大蛋白质)和物理化学性质的分子,这些分子与CPPs共价连接或非共价结合;(2) CPPs促进未结合的内体释放大分子物质(如环糊精和抗体);(3) 融合的脂质和肽改善CPPs的内体释放;(4)环状和其他构象受限的CPPs和CPMs的进入细胞溶质的效率提高。然而,是否所有CPPs /CPMs离开核内体或通过VBC机制转移穿过质仍有待确定。
细菌毒素
细菌毒素进入细胞的机制是多种多样的。一些毒素直接穿过质膜到达胞质溶胶;其他毒素则与细胞表面受体结合并通过内吞途径和内体逃逸进入细胞;还有一些毒素被运输到ER经逆行易位进入细胞质。然而,无论进入途径如何,毒素必须在到达细胞质之前的某个拓扑上穿过脂质双层。
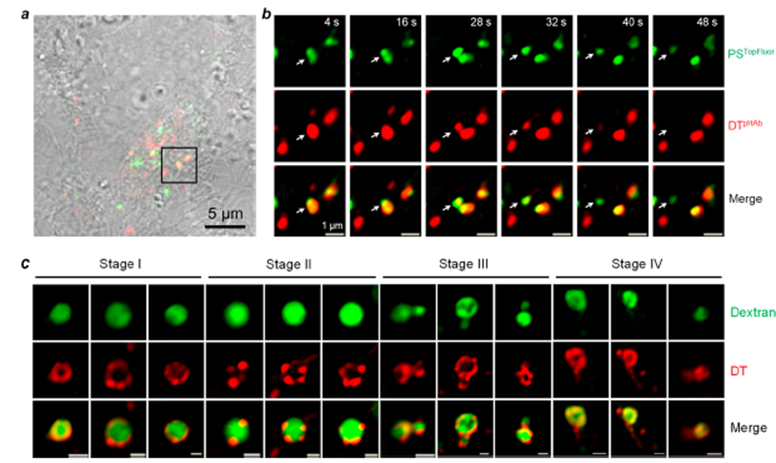
典型的哺乳动物细胞有数百个内体,它们通过不断的囊泡融合和裂变相互连接。核内体的横向融合加上毒素聚集成脂质结构域会逐渐将毒素分子集中到更少、更大的核内体中,最终达到“量子”VBC。这种情况为先前观察到的内体酸化和 DT 释放到胞质溶胶之间的浓度依赖性时间滞后提供了解释(即,在较低的DT 浓度下,滞后时间更长)。这也可以解释为什么在囊泡束缚和融合中起作用的 HOPS 复合物对 CPPs 体内的内体逃逸至关重要。
除细菌毒素外,还发现许多其他细菌和真核蛋白质可沿任一方向转移到质膜或内体膜上。例如,α-突触核蛋白的细胞间传递与帕金森病的进展有关;细胞渗透性自身抗体抗核DNA最初是在狼疮患者中发现的,这一发现导致了针对细胞内靶标的工程抗体疗法的开发。
病毒
病毒有助于揭示细胞中的基本机制(例如,RNA 剪接、自噬和细胞周期)。一般来讲,包膜病毒通过将膜与内体膜融合而逃离内体,而非包膜病毒必须破坏内体膜才能进入细胞质。已经发现了三大类“膜溶性”病毒因子:例如腺病毒蛋白VI,呼肠孤病毒的N-肉豆蔻酰化衣壳蛋白μ1和细小病毒VP1的磷脂酶A型2(PLA2)。三种病毒因子都通过诱导VBC来促进病毒内体逃逸。
一些研究小组最近通过使用先进的活细胞共聚焦显微学技术研究了多链体,脂复合物和LNP的内体逃逸。Rehman等人发现,在脂族和多链介导的siRNA递送过程中,核酸和载体都能从内体中突释出来,随后核酸在整个细胞质和细胞核中快速扩散。
已有实验证明,五种结构多样化的生物分子,包括线性CPP(Tat),环状CPP(CPP12),非肽CPM(CPM3),AB类细菌蛋白毒素(DT),和单结构域蛋白质毒素(NleC)是通过诱导VBC从内体膜逃逸到细胞质基质中的。此外,文献研究表明,其他细菌和真核蛋白以及非包膜病毒也可能通过VBC机制在任一方向上穿过细胞膜。通过解析VBC机制,我们现在可以合理化许多以前神秘且相互矛盾的观察结果。
参考文献:
Pei D. How Do Biomolecules Cross the Cell Membrane?[J]. Accounts of chemical research, 2022: 2601-2612.
推荐阅读 版权声明/免责声明

 点击这里,帮您找到理想的合作伙伴!
点击这里,帮您找到理想的合作伙伴!
本篇文章来源于微信公众号:药时代
发布者:药时代,转载请首先联系contact@drugtimes.cn获得授权
 为好文打赏 支持药时代 共创新未来!
为好文打赏 支持药时代 共创新未来! 