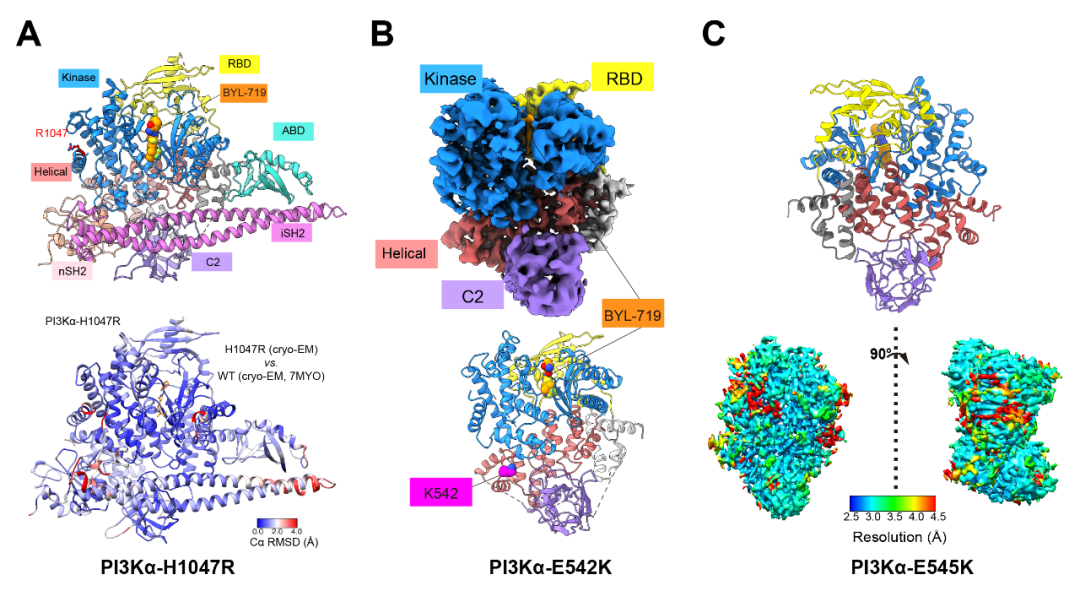
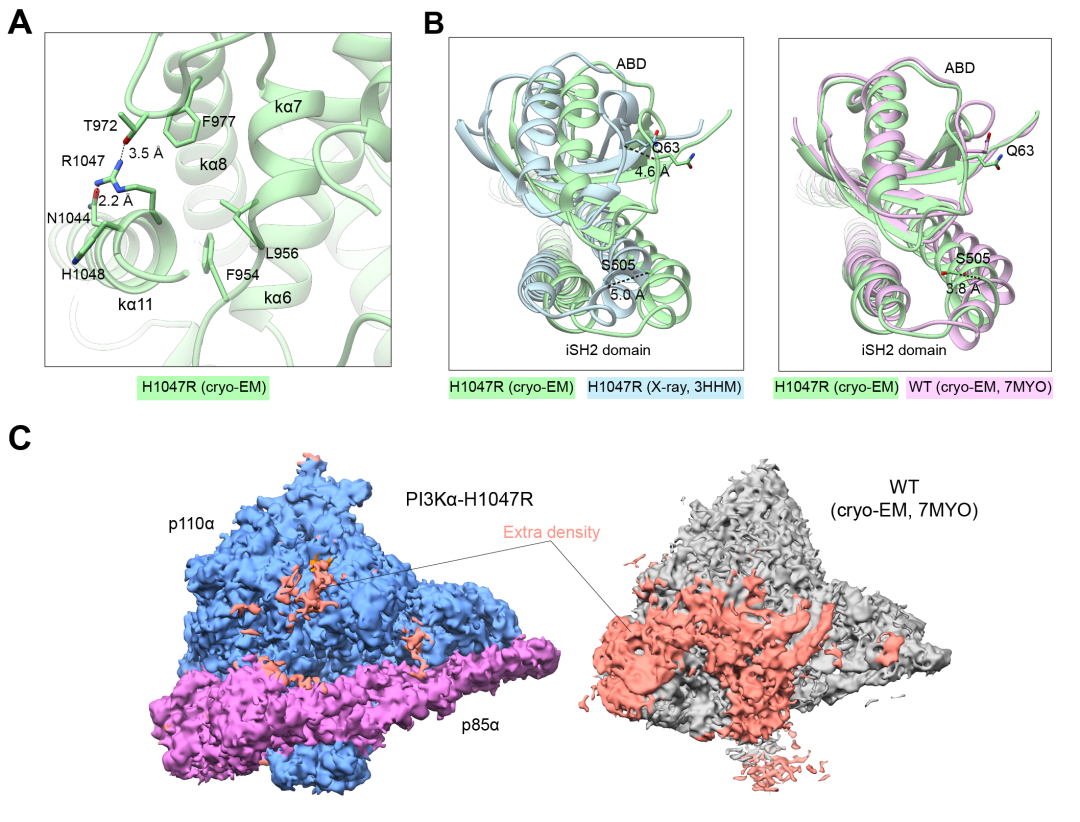
点击图片,预约本周四直播!2022年11月8日(北京时间),由复旦大学基础医学院王明伟、美国Scripps研究所Peter K. Vogt和中国科学院上海药物研究所杨德华领衔的合作团队于《美国国家科学院院刊》(Proceedings of the National Academy of Sciences of the United States of America, PNAS)在线发表了题为“Cryo-EM structures of cancer-specific helical and kinase domain mutations of PI3Kα”的研究成果。该论文首次报道了人源全长磷脂酰肌醇3激酶(Phosphatidylinositol 3-kinase, PI3K)α亚型(p110α-p85α)的三个致癌特异性突变体(H1047R、E542K和E545K)蛋白复合物的冷冻电镜结构,阐明了突变体诱导PI3Kα产生的局部和整体构象变化,为深入认识PI3Kα突变体致癌分子机制和创制特异性针对突变型PI3Kα的抗癌新药提供了重要的信息。
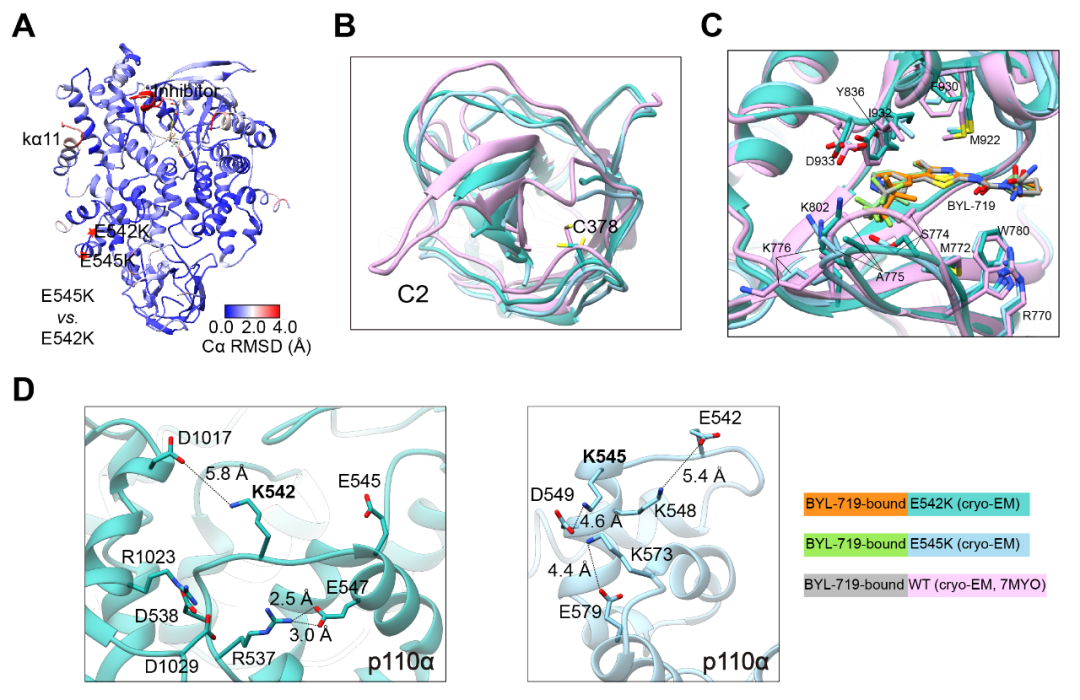
图3. A,突变体E542K和E545K模型的RMSD示意图(以Cα测量);B,突变体E542K、E545K和野生型(PDB ID:7MYO)结构6对比显示在C2结构域发生部分偏移;C,突变体E542K、E545K与野生型(PDB ID:7MYO)结构6中ATP结合位点的比较;D,突变体E542K(左)和E545K(右)在突变位点周围的氨基酸残基构象示意图。复旦大学基础医学院博士研究生刘晓和青年研究员周庆同为该论文的共同第一作者,复旦大学基础医学院王明伟讲座教授、美国科学院院士Peter K. Vogt教授和中国科学院上海药物研究所杨德华研究员为共同通讯作者。该研究先后获得了国家自然科学基金委员会、国家科学技术部、国家卫生健康委员会、中国科学院‒诺和诺德研究基金和美国国立卫生研究院的经费资助。据悉,两位审稿人对该文给予高度评价,认为其突破性地报道了三个热点突变体较之野生型结构的所有细节信息,是打开PI3Kα药物发现新篇章的奠基性工作,会被广泛引用。特别是p110α突变体的高精度结构解析将为设计高效选择性抑制剂提供关键路线图,是理解p110α致癌突变分子机制的重要线索。对于这项新突破,复旦大学附属肿瘤医院副院长、上海市胰腺肿瘤研究所所长虞先濬博士表示:PIK3CA编码PIK3α的催化亚基p110α,是一种突变率极高的致癌驱动基因,其中结直肠癌、胃癌、乳腺癌和脑癌中PIK3CA突变率高出25%,与患者较差的预后相关,是最重要的致癌基因之一。超过80%的PIK3CA突变出现在E542K、E545K和H1047R这三个热点位置上,是治疗恶性肿瘤的关键靶点。王明伟课题组及其合作团队首次解析了PIK3α三种致癌突变体复合物的三维结构,为理解PIK3CA突变致癌机理和靶向该激酶突变体设计及筛选特异性抑制剂提供了宝贵的结构基础,对临床干预PIK3CA活性和肿瘤精准治疗具有重要的意义。资深药物化学家、泰励生物科技有限公司联合创始人兼总裁仲伯禹博士认为:起始于PI3K的PI3K-AKT信号通路精准地调控着人体内细胞存活、代谢、生长和增殖等一系列生理功能。PI3K变异或者过表达所导致的过度活化能够引起抑瘤蛋白PTEN失活和多种癌症的发生,特别是PI3Kα亚型突变的致癌作用已经广为人知。然而,野生型PI3Kα关键性地调控着细胞的代谢过程,其活性一旦受到抑制,体内血糖和血酯将急剧升高,严重时会危及生命。长期以来,由于缺乏PI3Kα野生型和突变型的蛋白结构信息,人们无法选择性地抑制突变型PI3Kα之活性,而野生型被同时抑制而产生的毒副性严重阻碍了相关药物的研发。针对近年来PI3K抑制剂频频发生的临床不良事件及吉利德(Gilead Sciences)和因塞特(Incyte)对其已经获得加速批准后药物的撤回,数月前美国食品药品监督管理局(FDA)专门就PI3K抑制剂召开了肿瘤药物咨询委员会(ODAC)会议,要求未来的PI3K抑制剂在申请血液肿瘤适应症时应提供随机对照试验的数据支持,从而极大地提高了此类药物的创新门槛。王明伟/Vogt/杨德华领衔的合作团队首次解析了PI3Kα三种致癌突变体复合物的三维结构,不仅为研究PI3K的生理功能和突变致癌机制提供了新的理论基础,还为设计更安全的选择性地靶向突变型PI3Kα的新药分子贡献了极为有用的工具。长期困扰抗肿瘤药物研发领域的PI3K亚型选择性,尤其是α亚型中更具挑战性的野生型和突变型精准选择性药物将会在这项成果的启发下找到更快捷的解决方案,该工作的重要突破还能帮助研究人员迅速开发出更加安全高效的PI3Kα突变型高选择性药物,惠及相关肿瘤病人。冷冻电镜作为近年来涌现出来的颠覆性结构生物学方法,在蛋白质结构解析及与配体结合机理研究上发挥着越来越重要的作用。王明伟/杨德华团队不久前已在2型糖尿病治疗领域的胰高血糖素样肽-1受体(GLP-1R)、葡萄糖依赖性促胰岛素释放多肽受体(GIPR)和/或胰高血糖素受体(GCGR)多重激活机理的探索上获得了全球领先的进展,最近又在与肿瘤增殖密切相关的PI3K-AKT信号通路研究方面取得突破性斩获,不仅体现该团队在选择解决新药研发未满足需求上的卓越洞见和结构生物学功力,也彰显出他们对先进技术手段的快速掌握能力和高超的资源利用效率。文末点击“阅读原文”,查看论文原文!参考文献
1. Fruman D A, Chiu H, Hopkins B D, et al. The PI3K pathway in human disease. Cell, 2017, 170(4): 605-635.
2. Samuels Y, Wang Z, Bardelli A, et al. High frequency of mutations of the PIK3CA gene in human cancers. Science, 2004, 304(5670): 554-554.
3. Samuels Y, Diaz Jr L A, Schmidt-Kittler O, et al. Mutant PIK3CA promotes cell growth and invasion of human cancer cells. Cancer cell, 2005, 7(6): 561-573.
4. Kang S, Bader A G, Vogt P K. Phosphatidylinositol 3-kinase mutations identified in human cancer are oncogenic. Proceedings of the National Academy of Sciences, 2005, 102(3): 802-807.
5. Mandelker D, Gabelli S B, Schmidt-Kittler O, et al. A frequent kinase domain mutation that changes the interaction between PI3Kα and the membrane. Proceedings of the National Academy of Sciences, 2009, 106(40): 16996-17001.6. Liu X, Yang S, Hart J R, et al. Cryo-EM structures of PI3Kα reveal conformational changes during inhibition and activation. Proceedings of the National Academy of Sciences, 2021, 118(45): e2109327118.
7. Hart J R, Liu X, Pan C, et al. Nanobodies and chemical cross-links advance the structural and functional analysis of PI3Kα. Proceedings of the National Academy of Sciences, 2022, 119(38): e2210769119.













 为好文打赏 支持药时代 共创新未来!
为好文打赏 支持药时代 共创新未来! 