




全文共22873字,预计阅读时长为25分钟。
文章作者/来源:生信人 药时代授权转载,衷心感谢!
导览
人体微生物组成了一个复杂的多界群落,在多个身体部位与宿主共生互作。宿主-微生物的互作影响多种生理过程、是多种多因素疾病的条件。在过去10年,微生物群落被认为影响多种癌症的发生、发展、转移和对治疗的反应。然而微生物对癌症生物学存在影响的因果证据才刚刚被揭示,因此解析微生物对癌症的调节以及对癌症治疗产生影响在分子层面上的机制,具有重大的科学价值和临床意义。
在这篇综述中,研究者描述了不同促进癌症发生发展的微生物生态位共同具有的分子致病机制;强调了对相关问题理解的进步性、局限性、挑战和前瞻,这些问题包括微生物如何影响癌症和癌症对治疗的反应、微生物或其分泌的具有生物活性的代谢物如何具有潜在的利用价值、如何成为癌症治疗的精准靶标等等。
图表解读
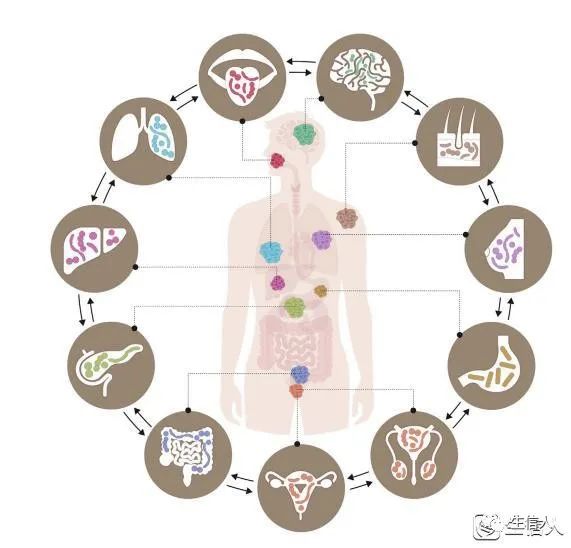
Fig1. 不同癌症类型的微生物生态位
微生物群落存在于多种黏膜表面,并且在多种癌症类型中以低生物量生态系统的形式存在。不同生态位之间的微生物组成可以多向共享。比如,原生于口腔的梭杆菌可以从口腔迁移,通过消化道或血液,沉积并定植在大肠。这种微生物群落被越来越多的研究者认为与癌症有关。
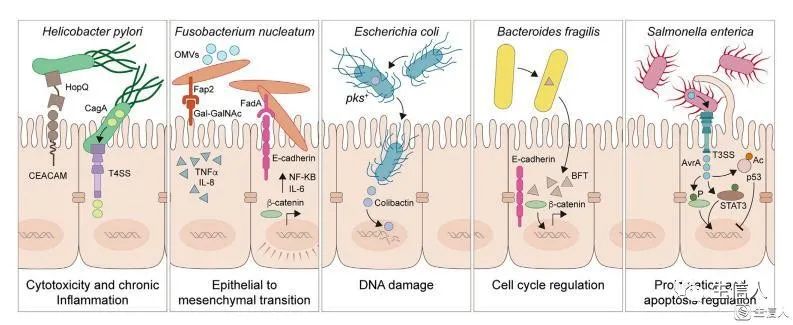
幽门螺杆菌 Helicobacter pylori – 细胞毒性和慢性炎症
幽门螺杆菌通过外膜黏附素HopQ结合CEACAM的方式与胃上皮细胞结合,毒力因子CagA通过T4SS直接注入上皮细胞。CagA激活Wnt/β-catenin通路导致细胞转换和凋亡失调。
具核梭杆菌 Fusobacterium nucleatum – 间充质转化
具核梭杆菌可能通过多种方式引发癌症。如毒力因子FadA黏附素使得细胞内化并诱导NF-κB和IL-6介导的促炎级联反应。另一种重要的黏附素Fap2在肿瘤表面与D-半乳糖-β(1-3)-N-乙酰-D-半乳糖胺(Gal-GalNAc)碳水化合物部分互作,通过Wnt/β-catenin通路增强细胞增殖、增加促炎细胞因子的产生、促进EMT,其中EMT是肿瘤细胞浸润、转移、干性和治疗耐受的突出特征。
大肠杆菌 Escherichia coli – DNA损伤
致病大肠杆菌具有一整套与致病性相关的毒力因子和毒素,包括分泌性遗传毒物colibactin。一旦被宿主细胞内化,colibactin会诱导DNA链间交联和双链DNA断裂,以及前肿瘤细胞转化和多种癌症基因组中检测到的突变特征。
脆弱拟杆菌 Bacteroides fragilis – 细胞周期调控
产肠毒素的脆弱拟杆菌(ETBF)编码BFT,是肠道细胞紧密连接的靶标,导致E-钙粘蛋白裂解。因此随着活性氧(reactive oxygen species, ROS)增加,通过NF-κB信号和组织损伤提高肠道通透性、引发慢性肠道炎症,最终导致CRC(结直肠癌)。
肠道沙门氏菌 Salmonella enterica – 增殖与凋亡调控
肠道沙门氏菌的3型分泌系统(T3SS)驱动毒力因子注入细胞内,比如多功能效应因子AvrA通过由Wnt/β-catenin通路引发的细胞增殖促进肿瘤形成。
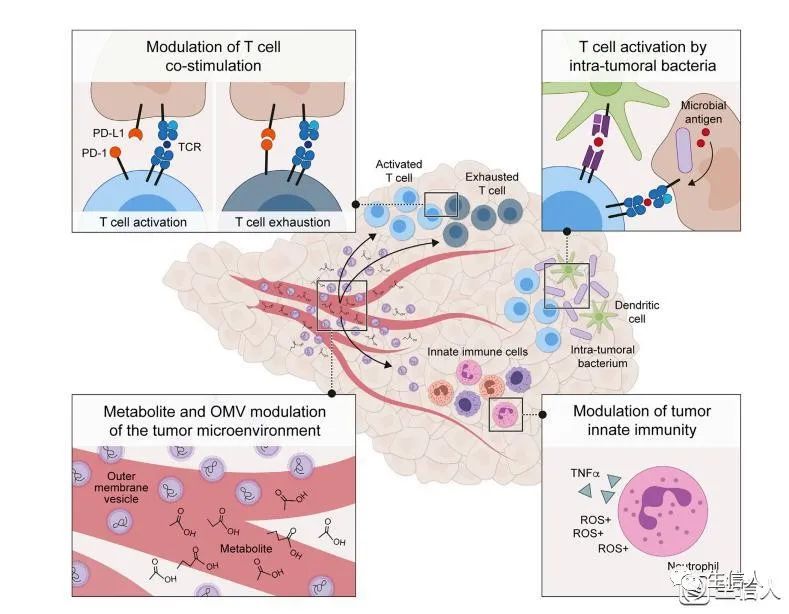
Fig3.肿瘤免疫微环境的微生物组调控
微生物分泌的代谢物、载物外膜囊泡(outer membrane vesicle, OMV)、肿瘤内细菌诱导一系列复杂的免疫调节作用,调节肿瘤生长、肿瘤内部及周围的免疫抑制环境。
微生物分泌的基团能够通过调控中性粒细胞等先天免疫细胞的活化和趋化,产生TNFα和活性氧(ROS+)对抗肿瘤细胞,影响肿瘤微环境(TME)的先天免疫反应。
微生物代谢物和OMV能够影响适应性免疫反应。通过调控T细胞的共刺激,它们能够影响肿瘤微环境的T细胞活化和衰竭,直接影响免疫检查抑制剂(immune checkpoint inhibitor, ICI)的疗效。
此外,细胞内细菌的肽片段能够直接呈递在肿瘤细胞表面,或通过HLA由专门的抗原呈递细胞呈递,从而驱动T细胞活化和潜在的癌症免疫反应。
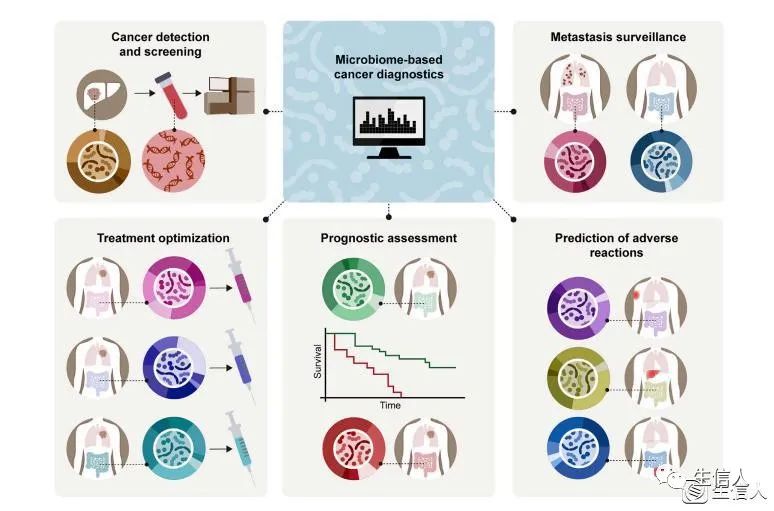
Fig4.微生物组数据在癌症诊断和患者分型中的应用
基于微生物组的癌症诊断:癌症的检测和筛查、转移监测、治疗优化、预后评估、不良反应的预测。
微生物组分析产生的数据可以促进新型癌症诊断能力的发展,包括通过鉴定外周血细菌DNA和RNA来检测癌症、测定癌症微转移的进展、评估预后、为个人定制治疗方案,以及利用人工智能算法预测患者的治疗反应和出现不良反应的风险。
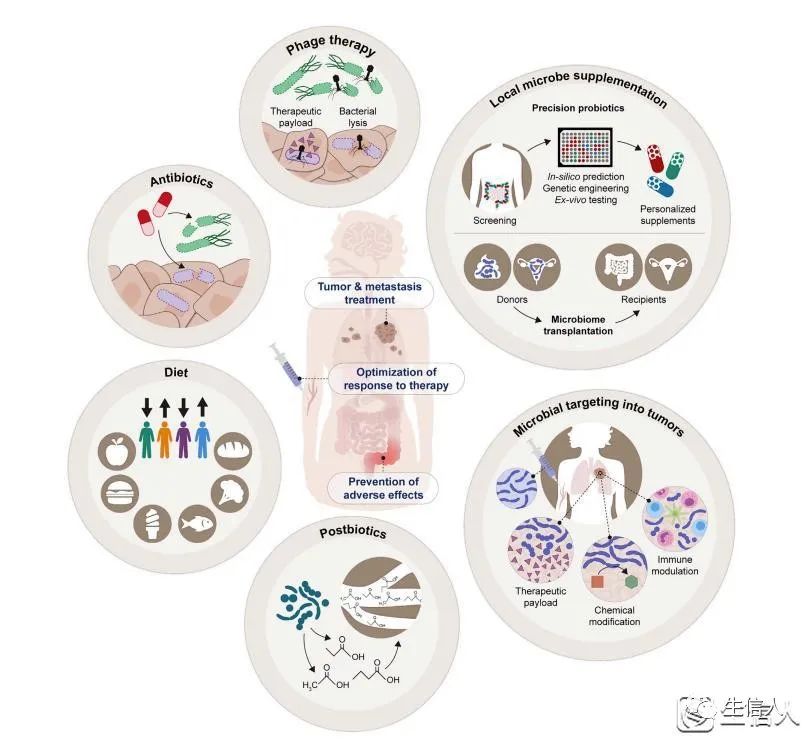
Fig5.癌症治疗中的微生物组调控
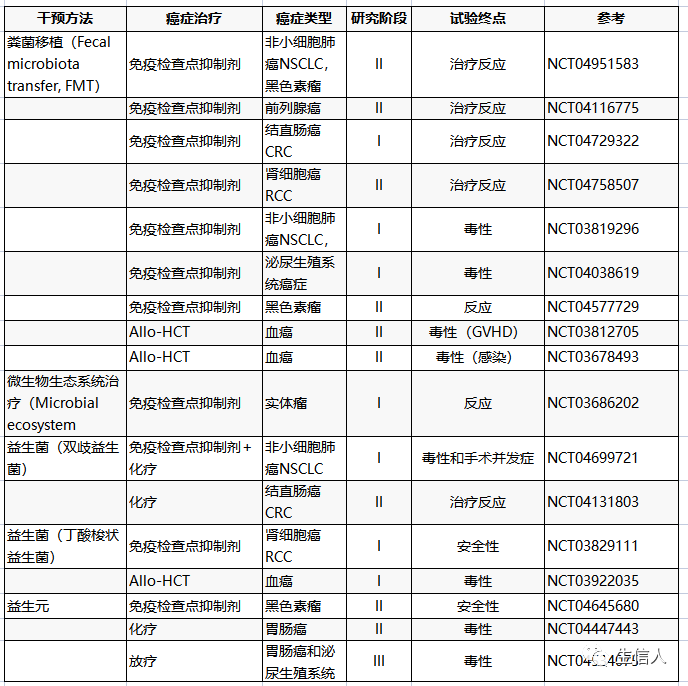
Table1.当前微生物组调节在癌症治疗中的作用的临床研究

据估计,人体内有3万亿细菌,它们协调着生理过程和疾病易感性的相互作用。虽然细菌细胞数目和人体细胞数目相当,但是细菌的遗传多样性比人体细胞高出100倍,其编码的基因能够使细菌具有很强的运动能力和代谢能力,不仅影响自身的生态位,还影响宿主组织特异性和免疫细胞的功能。除了细菌,人体微生物还包括真核生物真菌和原生动物以及病毒。总的来说,在健康的情况下,宿主与微生物共生,宿主提供营养丰富的微环境,微生物辅助消化和代谢。肠道微生物、皮肤微生物、口腔微生物的特征是数目多、种类丰富多样;阴道微生物也得到了同样充分的研究,但其特征是多样性低、高度特异且存在占优势的微生物类群。此外,一些器官和组织,如肺、前列腺、膀胱、乳腺、肝脏和胰腺曾经被认为是无菌的,但现在随着二代测序(next generation sequencing, NGS)的出现,这些器官和组织被认为存在潜在的低生物量的微生物类群。然而这些微生物是共生的位置特异性种群,还是暂时从相邻位置迁移来的仍有待商榷。
癌症的发展和进展大体上被认为多数具有可区分的标志物,但这些标志物有时需要互相补充,有时会出现重合。通过持续增殖、避免细胞生长受限、激活侵袭和转移途径、启动重复免疫、促进新生血管生成、抵抗自噬等方式,癌细胞能够有效增殖并实现免疫逃逸。这些过程已经被广泛地研究了数十年,但微生物组对癌症发生、发展、治疗反应的潜在影响仍不清楚。在疾病的情况下,每个生态位的微生物都可能影响癌症的发展,可能通过微生物组成变化引发的群落水平的互作,也就是术语生态失调(dysbiosis);也可能是通过单独种群的直接作用;也可能是通过分泌的或调节的分泌物。特定生态位的微生物组能够影响癌症,如口腔中的微生物组能够调节口腔癌症(口腔鳞状细胞癌[oral squamous cell carcinomas, OSCC])、结肠癌症(结直肠癌[colorectal cancer, CRC])、胰腺癌(胰腺导管腺癌[pancreatic ductal adenocarcinoma, PDAC])。男性和女性盆腔内器官的微生物特征对泌尿系统癌症和妇科癌症有重要的指示作用,包括前列腺、子宫颈、子宫内膜、卵巢和膀胱等部位的癌症。此外,“肿瘤内微生物”可能会更加促进局部肿瘤的发生。
研究癌症相关微生物组
微生物组组成和功能的特征能够通过多种方法的组合来确定,各种方法能够从不同方面理解这个复杂的群落结构。在认识到这些方法、模式的优势和局限后,人们可以越来越多地整合微生物大数据,定义人类癌症的生理学、病理学关联,同时尽量减少偏差和错误。
第二代测序
16S rRNA基因测序(16S rRNA gene sequencing)是较为广泛使用、价格低廉的NGS平台,能够高通量平行测定微生物组的群落结构。但是,在16S rRNA基因扩增区、数据整理和参考数据库等方面的放大偏差、嵌合体产生、方法学差异等问题,给分析输出数据、进行数据解释和跨研究的可重复性带来了挑战。
鸟枪法宏基因组测序(shotgun metagenome sequencing)不仅可以确定样本组分的分类,还能够评估基因,包括那些能够编码代谢途径的基因,以描述共生微生物群落基因组功能的潜力。鸟枪法宏基因组测序不加选择地对整个基因组进行测序,要求更高的测序深度和更高的花费,相比16S rRNA基因测序能够提供种水平甚至菌株水平的比对结果。然而研究之间的组分可重复性仍然是鸟枪法宏基因组测序中的挑战,这种方式的功能输出能够提高研究之间的一致性。
宏基因组测序能够利用序列库在计算层面上重构微生物群落的组成,从而在尚未命名的物种中识别新的微生物基因组。这些尚未被培养方法或宏基因组学鉴定的微生物组成分,即所谓的微生物“暗物质”,可能与癌症研究特别相关,因为肿瘤在宿主内表现出高度非生理环境。
宏转录组学(metatranscriptomics)利用NGS能够描述单种细菌或整个微生物组的转录组图谱,是衡量微生物活动和功能更直接的方式,同时能够衔接宏基因组和下游代谢组以及蛋白质组的读数结果。
低生物量肿瘤微生物的评估
为了精确描述低生物量肿瘤内微生物组,这种NGS形式试图解释背景噪音和污染。肿瘤组织的多区域16S rRNA基因测序显示,肿瘤实体间肿瘤内微生物群落的多样性和组成存在显著差异。这个方法的流程包括严格的组织处理、DNA提取、多重试剂盒组相关(‘‘kitome’’-related)、扩增控制,最终移除94.3%被推定为污染物的细菌。另一种方法是,利用肿瘤组织活检产生的鸟枪法测序数据描述癌症遗传学特征,挖掘探索肿瘤内的微生物组。TCGA测序数据分析显示,肿瘤和病人血液中的微生物特征可以被用来区别癌症亚型。然而,由于缺少对样本处理和测序的控制,区分内源性细菌和污染细菌只能通过生物信息去污算法实现(bioinformatic decontamination algorithms)。即使采用芯片去污( in silico decontamination )也能移除大量被推断为污染物的分类单元。鉴于这些技术和分析方面的挑战,采用多种方法进行研究越来越普遍,方法包括16S rRNA基因测序、多重荧光原位杂交、宏基因组学以及代谢组学等,显著提高低生物量癌症微生物组评估的准确性和稳健性。
非基因组的微生物组特征
代谢组学(metabolomics)是一种基于质谱分析的方法,描述由不同微生物产生或调节的数千种潜在的生物活性代谢物。利用这种方法,能够通过短链脂肪酸(short-chain fatty acids, SCFAs),对使用抗PD -1型免疫检查点抑制剂治疗的癌症患者的无进展生存进行分层,这些短链脂肪酸主要是乙酸、丙酸或丁酸等微生物降解淀粉的主要代谢物。
宏蛋白质组学(metaproteomics)描述与微生物组组成相关的数千种肽段和蛋白质,但是目前为止很少被应用,它在癌症研究中的潜力仍有待确定。
主要组织相容性复合体(major histocompatibility complex, MHC)的多肽组学(peptidomics)能够描述肿瘤细胞或免疫细胞上表达的基于蛋白质的多肽,这些肽段能够调节针对肿瘤的局部和全身的免疫应答。这种方法鉴定到黑色素瘤肿瘤内细菌相关肽段,这些肽段由人类白细胞抗原(human leukocyte antigen, HLA)呈递在癌症细胞表面,具有在肿瘤微环境中局部激活免疫应答的潜力。
除了高通量测序,多条件微生物组培养(培养组,culturomics)大大增加了可培养共生菌的库,使得在微生物群落中对单种细菌进行精确分类和功能评估成为可能。例如这种培养组大规模筛选显示,市场上24%的非抗生素药物抑制人体肠道细菌的生长。
肿瘤-微生物组分析
在癌症-微生物组研究中,解释和分析以上的多维大数据集,并把结果整合到生理层面仍然是在技术和概念上都十分艰巨的挑战。从技术上来说,这种大数据的整合十分重要,因为实验室内部的方法应用于同一数据集上都可能导致不一致的结论。从概念上来说,只有少部分推定的具有驱动作用的改变(driver changes)能够表明共生菌和相关功能是在癌症形成、发展和治疗反应之前促进其发生的,然而与癌症相关的微生物通常是被动的改变(passenger changes),是继发于癌症期间发生的生理变化的。由于测序和数据采集方法具有多样性,将因果关系从相关性中分离出来是该领域面临的巨大挑战。机器学习和其他基于人工智能的方式可能会更好地识别临床相关性中的潜在因果关系。通过独立发现和验证队列,这些方法流程可能有助于识别疾病或性状相关的微生物特征,同时也会考虑微生物组数据中的个体间差异。值得注意的是,机器学习能够识别复杂的大数据集中潜在的因果关系特征,但是仍旧需要实验验证来充分说明因果关系的确定性。尽管存在局限性,我们仍应在未来的癌症-微生物组研究中越来越多地使用人工智能方法。重要的是,对癌症患者在患病和治疗过程中随时间变化的纵向微生物组数据的需求在增加,就需要调整分析方法,以解决评估微生物组随时间动态变化中的难题。
癌症促进微生物
目前,只有11种生物(7种病毒、3种扁形动物和1种细菌)被确认为人类癌症的确切病因。这11种生物分别是人类疱疹病毒4型(Epstein-Barr, EBV)、乙肝病毒(Hepatitis B, HBV)、丙肝病毒(Hepatitis C, HCV)、卡波氏肉瘤疱疹病毒(Kaposi sarcoma herpesvirus, KSV)、人类免疫缺陷病毒(human immunodeficiency virus-1, HIV)、人乳头状瘤病毒(human papilloma viruses, HPV)、Ⅰ型人类T淋巴细胞病毒(human T cell lymphotropic virus type 1, HTLV)、肝吸虫(Opisthorchis viverrini)、(Clonorchis sinensis)、华支睾吸虫Schistosoma haematobium)、幽门螺杆菌(Helicobacter pylor)。这些微生物都通过多种机制促进癌症发展,包括诱导B细胞分化、中断细胞周期、免疫过度活化(在EBV、HBV、HCV和HIV感染中)、T细胞调节异常(在EBV和HTLV感染中)等,肝炎病毒和KSV能够直接诱导肝癌和卡波西肉瘤的发生。KSV和HPV都能直接通过致癌蛋白有效减少细胞凋亡。三种致癌扁形动物与胆管癌和肝癌密切相关,其中华支睾吸虫(S. haematobium)还与膀胱癌有关,扁形动物能够引发慢性炎症导致氧化应激反应和DNA毒性。
癌症相关的细菌
尽管微生物和微生物组的研究不断发展,国际癌症研究机构(International Agency for Research on Cancer, IARC)的致癌微生物列表在最近十年都没有更新。但是最近的的研究显示,除了幽门螺杆菌,还有数十种微生物能够调控或促进癌症发生。
Helicobacter pylori
幽门螺杆菌是公认的致癌物,年龄标化发病率(age-standardized incidence rate)为每年每10万人8.7例,世界半数人口中都能检测到幽门螺杆菌。总体来说,幽门螺杆菌通过干扰Wnt/b-catenin通路调节细胞转换和细胞凋亡,促成消化性溃疡、胃癌和黏膜相关淋巴组织(mucosa-associated lymphoid tissue, MALT)淋巴瘤。幽门螺杆菌能直接影响癌症发展,如Correa通路所述,多形核中性粒细胞的募集和单核淋巴细胞引发慢性炎症反应,通过白介素-1B(IL-1B)、肿瘤坏死因子α(TNFα)、干扰素γ(IFNγ)的促炎信号刺激主要的Th1型反应。幽门螺杆菌通过外膜黏附素 HopQ结合癌胚抗原相关细胞黏附分子(carcinoembryonic antigen-related cell adhesion molecules, CEACAM)附着于胃黏膜上皮细胞,由此,CagA可以通过4型分泌系统(type 4 secretion system, T4SS)直接注入上皮细胞。在胃黏膜上皮中,幽门螺杆菌与SHP2磷酸酶和PI3K相互作用,提示幽门螺杆菌直接参与上皮细胞向肿瘤细胞转化。
Fusobacterium nucleatum
在探究结直肠肿瘤中是否存在细菌的研究中,Castellarin等人发现在CRC肿瘤组织中具核梭杆菌转录本是正常组织的400倍,这提示与OSCC相关的口腔病原微生物可能影响远处身体部位的癌症。此外,具核梭杆菌与肝转移有关,扩大了其在癌症中的潜在影响。具核梭杆菌通过FadA黏附素附着于宿主的上皮细胞和内皮细胞,使细胞内化病原体,引发由NF-κB和IL-6街道的促炎级联反应,这可能是具核梭杆菌侵袭OSCC细胞的方式。在体外,产生外膜囊泡(outer membrane vesicle, OMV)的具核梭杆菌通过结直肠上皮细胞分泌IL-8和TNFα引发炎症。此外,具核梭杆菌通过Fap2半乳糖结合凝集素与CRC和乳腺癌表面的D-半乳糖-β(1-3)-N-乙酰-D-半乳糖胺(Gal-GalNAc)碳水化合物部分互作,定位于肿瘤位点。具核梭杆菌还有助于上皮细胞的间充质转化(epithelialmesenchymal transition, EMT),EMT是癌症细胞侵袭、转移、干性和治疗抗性的主要特征。
Escherichia coli
大肠杆菌通过诱导炎症、氧化应激、细胞生态位改变以及对宿主细胞周期的干扰和操控,促进癌症的形成。由致病性大肠杆菌pks毒力岛编码、分泌的遗传毒物colibactin,能够诱导真核生物的上皮细胞DNA链间交联和双链DNA断裂。此外,在CRC小鼠模型和CRC人体活检中发现,大肠杆菌可能通过产生促进细胞增殖和肿瘤生长的生长因子加速衰老。值得注意的是,大肠杆菌诱导的DNA损伤反应和对Wnt信号的间接影响(这两个过程都促进细胞性状转化)在CRC中都很常见。
Bacteroides fragilis
脆弱拟杆菌是活跃于整段结肠的共生细菌,其中产肠毒素的脆弱拟杆菌(enterotoxigenic B. fragilis, ETBF)由于在癌症患者的粪便和黏膜样本中富集,被认为与诱导结肠炎和结肠肿瘤的发生有关。ETBF编码金属蛋白酶脆弱拟杆菌毒素(Bacteroides Fragilis Toxin, BFT),能够在CRC中,通过靶向肠道细胞的紧密连接、剪切E-钙粘蛋白诱导慢性肠道炎症和组织损伤,增加炎症和细胞通透性。此外,Wnt/β-catenin、NF-κB信号通路以及Th17适应性免疫也在这个过程中被激活。从机制上来说,感染ETBF可能会通过上调依赖toll样受体(toll-like receptor, TLR4)途径的表观遗传和转录调节水平,起到细胞干性调节的作用,在体内和体外都能促进结直肠癌的发生。
Salmonella enterica
在感染沙门氏菌的小鼠模型和细胞中,MAPK和AKT途径的活化对胆囊癌维持细胞性状转化至关重要。3型分泌系统能够转入靶细胞效应蛋白,如类cyclomodulin蛋白伤寒毒素(cyclomodulin-like protein Typhoid toxin)和多功能效应因子AvrA(multifunctional effector AvrA),参与遗传毒物介导的突变引发的癌症。此外,类cyclomodulin蛋白使得细胞内细菌生存且促成细胞失调,AvrA促进Wnt/β-catenin和JAK/STAT通路激活、促进细胞增殖和分化、增强靶向p53的乙酰转移酶的活性,二者共同驱动细胞周期阻滞、抑制凋亡,导致肿瘤发生。
癌症相关的非细菌微生物
微生物组中除细菌外的其他界,如病毒组(virome)、真菌组(mycobiome)和寄生组(parasitome)也可能导致癌症。
病毒
病毒组由98%的噬菌体和2%的真核细胞病毒组成,与细菌微生物组相似的是,病毒组同样复杂,存在于身体多个部位,既有共生病毒也有致病病毒,且病毒组在出生1个月后就形成了。最近的研究表明整体的病毒组特征可以作为癌症进展的标志物。肺腺癌以及口腔、乳腺、结肠、泌尿生殖系统中超过100种癌症的宏病毒组特征已经被检测。Cantalupo等人认为HPV除了作为宫颈癌的确定病因和其他泌尿生殖系统的潜在病因外,还与头颈癌、膀胱癌的进展有关。在这些病例中,基因的表达谱和突变谱显示HPV可能驱动肿瘤发生,但是HPV参与诱发癌症的确切机制仍不清楚。
除真核细胞病毒外,噬菌体也可能作为癌症的调节因子。例如,一些噬菌体可能与癌症细胞互作,下调整合素和其他参与致癌作用及转移的蛋白。Syphoviridae和Myoviridae科的其他噬菌体在CRC患者的粪便中富集。因为这些噬菌体多数是温和噬菌体,因此尚不清楚这些结果是真实的生物信号,还是参考基因数据库中温和噬菌体过表达。另外与对照组相比,CRC患者的肠道中检测到多种链球菌特异性噬菌体和一种弧菌噬菌体增加。因此推断这些噬菌体的改变可能引起基因交换,建立起宿主内的生态适应(ecological adaptations)和群落网络(community networking),进而影响癌症。尽管存在这些关联,但噬菌体对癌症发生的直接影响仍未被证明。病毒、病毒相关细菌、真核细胞之间跨界相互作用可能与癌症相关的很好例子是,皮肤相关的表皮葡萄球菌能够激活皮肤内源性逆转录病毒的表达,进而引发共生菌特异性的T细胞应答,促进组织修复。类似的微生物机制是否会影响癌症相关过程可能是未来数年癌症-微生物组研究的重要方向。
真菌
真菌组失调越来越多地被认为与多种疾病有关,包括急性移植物抗宿主病(acute graft versus host disease)、口腔癌和CRC。14个真菌标志物与CRC有关,同时与菌群失调有关,比如在结直肠癌中富集梭杆菌门(Fusobacteria)、在胰腺癌中富集马拉色霉菌属(Malassezia)。从机制上来说,马拉色霉菌迁移到胰腺,通过真菌细胞壁结合宿主的甘露糖结合凝集素(mannose-biding lectins)激活补体级联反应。最近也有研究结果显示白色念珠菌(Candida albicans)与胃癌有关,甚至可以作为胃癌的预测因子。几种推定的机制可能可以解释白色念珠菌和癌症风险的关系,比如可以产生已知能够改变口腔癌细胞增殖的亚硝胺;通过TNFα和IL-18诱导免疫调控,促进肿瘤细胞黏附上皮细胞。这些都值得未来进行机制研究。不管健康还是患病的情况下,真菌组和细菌组可能都会相互影响,同时也会和宿主互作;例如使用抗生素后真菌的恢复,另外真菌-细菌互作也会在乳腺癌和黑色素瘤小鼠模型进行放疗时驱动不同的肿瘤反应。当然,很多真菌组对癌症潜在的因果影响在机制上还不清楚。
寄生虫
除国际癌症研究机构(IARC)确认的寄生致癌物外,寄生组对癌症发展的影响还未被完全揭示。寄生微生物可能通过诱导促进癌症的菌群失调间接调控其他微生物组。例如,被华支睾吸虫(S. haematobium)感染的膀胱癌患者中,沙门氏菌(Salmonella)、克雷伯菌(Klebsiella)和大肠杆菌(E. coli)等革兰氏阴性菌更为多见。此外,寄生微生物可能通过诱导免疫调控直接调控癌症发展。例如,多种寄生微生物都能通过调节TLRs和炎症小体、诱导调节性T细胞活化等方式诱导先天性免疫和适应性免疫。除了促进癌症发生,寄生微生物还可能有益于抗癌的一些生理活动,甚至可以作为癌症治疗的佐剂。以上提到的和未提到的寄生微生物对癌症的影响都值得进一步的机制探究。
微生物群落的改变与癌症
微生物组中除了单独的致病微生物或共生微生物直接影响癌症发展外,共生生物中细菌整体的紊乱更多地被认为与癌症发展有关,即所谓的菌群失调(dysbiosis),菌群失调可能通过包括细菌-细菌互作等多种群落驱动的机制促进癌症发展。以口腔为例,吸烟和饮食变化引起特征微生物种群水平上的改变,导致生态位的改变(如pH改变),进而影响共生微生物的种类以及共生微生物分泌的副产物。口腔菌群失调已经被认为与龋齿、牙周炎和口咽癌(如OSCC)的发生有关,这种失调部分由变异链球菌(Streptococcus mutans)、具核梭杆菌(F. nucleatum)和牙龈卟啉单胞菌(P. gingivalis)等条件致病菌介导。类似地,下消化道微生物组的改变与消化道癌症和CRC有关。例如,健康的肠道微生物通常由乳酸杆菌(Lactobacilli)、拟杆菌(Bacteroides)和双歧杆菌(Bifidobacterium)等优势种群组成,而CRC中则是梭杆菌(Fusobacterium)、卟啉单胞菌(Porphyromonas)、Parvimonas、消化链球菌(Peptostreptococcus)和孪生球菌(Gemella)过度优势,说明发生了菌群失调。
如果考虑到肿瘤发生的不同阶段,这种菌群失调的相关性会更加复杂。Nakatsu等人报道结直肠肿瘤发生过程中,即从腺瘤到癌症的过程中,每个阶段都有独特的微生物群落结构形成的微环境。相比邻近的黏膜,结直肠病变位点具有疾病特异性的微生物群落结构,比如梭杆菌门(Fusobacteria)等口腔相关的微生物更为优势。另一个例子是,健康的阴道微生物组以乳杆菌属(Lactobacillus)为优势共生菌,具有多样性较低的特征。有结果显示,厌氧菌会导致细菌性阴道病(bacterial vaginosis, BV),而白色念珠菌(Candida albicans)等真菌的增加可能导致以局部炎症反应为特征的阴道炎。除HPV外,BV也与宫颈癌前病变和上皮内瘤变有关。最近的一项研究确认BV是HPV阳性的宫颈癌患者的共同作用因素,表明菌群失调在引发宫颈癌中起直接作用或辅助作用。值得注意的是,以上菌群失调与癌症的关系多数只能是相关关系,而不是因果关系。因此,还不能确定微生物种群的改变是造成癌症发生的原因还是肿瘤出现的结果。区分因果将是未来十年癌症-微生物组研究的重点。
肿瘤内微生物
除了黏膜生态位(消化道、呼吸道、泌尿生殖道和皮肤)的微生物,最近有研究者提出肿瘤内微生物(intratumoral microbes 或 tumor microbiome)可能构成独特的低生物量的生态系统。肿瘤内微生物可能影响肿瘤微环境,包括炎症介质,如组织驻留和外周招募的免疫细胞(髓样细胞、T细胞、B细胞和NK细胞)、纤维母细胞、内皮细胞、脂肪细胞和周细胞。7种癌症类型超千个肿瘤样本和邻近非肿瘤组织的分析确定了肿瘤类型特异性的微生物组由大多数肿瘤内细菌组成。例如,肺癌和胰腺癌中发现了肺炎克雷伯菌(Klebsiella pneumoniae),胰腺癌和乳腺癌中存在阴沟肠杆菌(Enterobacter cloacae)、弗氏柠檬酸杆菌(Citrobacter freundi)、肠杆菌(Enterobacter asburia)、梭杆菌(Fusobacterium)等。这些微生物是组成了预先确定的生态位,还是只是暂时的随机定植还不确定。例如,胰腺内肿瘤微环境发现的与癌症相关的微生物可能会通过围肠易位穿过胰腺管到达胰腺。肠道上皮屏障损伤可能会影响肿瘤内微生物种群,因为微生物可能会随血液扩散在肿瘤组织定植。最近,由研究表明,CRC相关的肿瘤内大肠杆菌在肠道血管屏障受损后迁移到肝脏,通过招募先天免疫细胞和炎症免疫细胞改变肝脏微环境直接促进癌症转移。此外,在癌症发展过程中,宫颈中增多的致癌细菌可能到达且定植在子宫和卵巢。
这些肿瘤内微生物对肿瘤微环境的作用和相关致癌机理的研究才刚刚开始,值得对机制进一步探索。预测模型显示肿瘤中出现的细菌可能在特定身体部位上调特定的途径。例如,在肺癌中富集的细菌可能具有代谢香烟相关代谢物的潜在能力。在PDAC中,肿瘤内微生物通过丙型变形菌纲的细菌( Gammaproteo)产生的胞嘧啶脱氨酶的特异变体使化疗药物吉西他滨(gemcitabine)失活。不管利用无菌小鼠还是使用抗生素去除微生物,都能促进肿瘤微环境中免疫反应的重编程,减少髓系抑制细胞(myeloid-derived suppressor cell, MDSC)的渗透,增加CD4+ Th1细胞极化,激活CD8+ 细胞毒性T细胞,从而显著减少癌变,增加免疫治疗的疗效。
微生物影响癌症的机制
多种接触依赖、非接触依赖、免疫机制驱动错综复杂的宿主-微生物相互作用,使得微生物组能够诱导癌症的调控。值得注意的是,单种微生物通过多效相互作用对肿瘤发生产生影响,这些相互作用可能是借助毒力因子的表达和分泌、物理结合诱导的信号、免疫细胞的招募等共同促进致癌影响。理解其中的机制对于把这些相互作用作为癌症诊断标志和治疗方法至关重要。
接触依赖的互作
许多固有共生微生物和侵入性病原体可能通过直接与宿主细胞相互作用影响癌症相关过程。例如,在胃黏膜中,幽门螺杆菌(H. pylori)将CagA注入到上皮细胞中,CagA直接与E-钙粘蛋白互作,扰乱E-钙粘蛋白/β-连环蛋白的相互作用,使得细胞核β-连环蛋白聚集,并最终诱导下游的肠道差异标志分子表达,导致肠道癌症恶化前的组织变化。相似的是,肠道沙门噬菌(S. enterica)注射AvrA到细胞内,上调β-连环蛋白途径,激活STAT3信号、炎症反应以及间充质转化诱导的转录因子。聚核梭杆菌(F. nucleatum)与E-钙粘蛋白之间直接的相互作用参与和FadA的结合,这一过程通过增加下游E-钙粘蛋白/β-连环蛋白调控的转录因子的表达,导致DNA损伤、上皮细胞增殖、细胞干性的获得以及细胞极性的丧失。细菌还能够通过接触依赖释放不同种类的cyclomodulin,激活宿主细胞内活性氧和活性氮的释放,对宿主细胞周期产生影响。细胞致死膨胀毒素(cytolethal distending toxin, CDT)是由大肠杆菌、螺杆菌和沙门氏菌产生的一种cyclomodulin,能够与宿主细胞上的一种未知受体结合,引起细胞内化,并最终通过DNA损伤抑制细胞增殖。这些细胞周期和DNA损伤的调控可能有利于癌症的发生。
非接触依赖的互作
微生物能够产生、代谢生物活性分子和外膜囊泡(OMV),这些物质通过体循环到达癌症位点以及癌症转移位点,从而远程、间接地影响癌症发生。分泌的代谢物、发酵产物、分解产物在不同肿瘤位点都具有活性。例如,脂多糖(LPS)毒素和乙酸盐能够通过促进间充质转化(EMT)以及血管生成促进癌症发生;而戊二胺能够在体外逆转EMT,抑制细胞移动和侵袭。宿主分泌的代谢物能够接着被微生物代谢,比如次级胆汁酸(secondary bile acids, sBAs)、脱氧胆酸(deoxycholic acid, DCA)和石胆酸(lithocholic acid, LCA),与CRC、肝细胞癌等癌症的发生有关。CRC活检与临近的健康组织相比,具有更高浓度的硫化氢,硫化氢是微生物产生的具有遗传毒性的化合物。在体外,硫化氢调节细胞能量代谢和细胞增殖。微生物调节的代谢物可以来源于饮食,比如在克罗恩病中,脂肪性的饮食会增加胆汁酸,比如梭菌属产生的Tyr-Chol,可能会在临床背景下促进炎症诱导的癌症发生。另一方面,在小鼠中,胆汁酸被共生细菌代谢为次级胆汁酸,通过诱导CXCR6+自然杀伤T(NKT)细胞应答促进皮下肿瘤的发生和肝转移。
在某些情况下,微生物调控的代谢物对癌症发展的影响可能是生态位特异性的。Kadosh等人最近发现,一种微生物来源的没食子酸(gallic acid)参与了不同肠道位点的p53变异,其造成的结果也不同,会导致近端肠道的肿瘤抑制或远端肠道的恶性细胞转化。微生物组也能够代谢宿主激素影响癌症发生。例如,包括大肠杆菌和肠杆菌其他种等共生细菌能够使雌激素代谢物解离,解离的代谢物以具有生物活性的形式通过肝肠循环(enterohepatic circulation)进入宿主的门脉循环(portal circulation),促进雌激素介导的癌症,如乳腺癌和前列腺癌的发生。在男性个体中,肠道和尿道的微生物能够通过产生糖皮质激素氧化雄激素的中间产物增加前列腺癌的风险。
细菌胞外外膜囊泡(OMV)是参与癌症发生过程的重要细菌传递系统,OMV主要由革兰氏阴性菌产生。OMV使得细菌能够传递遗传物质、免疫调节分子、毒力因子和毒素到宿主循环系统中,其细胞膜的封装可以作为抗体和防御素的诱饵。重要的是,包括CagA、VacA、BFT等致癌分子在内的许多毒力因子,以及大肠杆菌分泌的遗传毒物colibactin,能够通过OMV运输促进远端部位的肿瘤发展。例如,携带CagA的幽门螺杆菌OMV能够改变核小体内DNA与组蛋白的结合、破坏上皮细胞的结合,导致细胞转变为具有侵袭性的间充质细胞,进而发展为癌症。
此外,黏膜部位的共生微生物能够通过对抗病原菌和病原体的定植和入侵间接影响微生物对癌症的作用。这种共生微生物和宿主的相互作用发生在身体的不同位点,通过黏膜屏障诱导定植抵抗、调节黏膜抗病原的免疫反应、以及酸性、氧化作用和铁利用率等生物物理特性的改变来实现相互作用的发生。
免疫互作
癌症中免疫系统-微生物的相互作用要么发生在黏膜表面,通过微生物代谢物和外膜囊泡影响全身,要么在淋巴器官内或肿瘤微环境中产生局部影响。局部和远端的微生物信号能够影响先天性免疫反应和适应性免疫反应,导致系统性的或肿瘤微环境内的免疫调节以及免疫监视。微生物产生的代谢物能够通过体循环到达远端的肿瘤实体,激活抗肿瘤或促进肿瘤发生的先天免疫反应。例如,来自共生微生物或病原体的进化保守的微生物相关分子模式(microbe-associated molecular patterns, MAMPs)可能会被先天性免疫系统通过TLR、NOD样受体等模式识别受体(pattern recognition receptors, PRRs)感知,引发先天性免疫应答和适应性免疫应答的先天性免疫部分。细菌MAMPs能够通过增强TLR信号、作为癌症疫苗佐剂增强抗肿瘤免疫。还有肠道微生物调节的次级胆汁酸(sBAs)等生物活性代谢物影响针对肿瘤的先天性免疫,通过调节肝细胞癌肿瘤微环境中的肝脏NKT细胞控制肿瘤进展。
微生物产生的信号能够改变肿瘤相关的适应性免疫反应。肿瘤微环境中T细胞衰竭是免疫系统未能清除肿瘤细胞的重要现象,这种现象由T细胞表达的芳烃受体调控。这个受体由5-羟色胺激活,而5-羟色胺是肠道微生物诱导的色氨酸代谢的副产物,从而导致微生物促进免疫介导的肿瘤发生的结果。相反的,定植在回肠的肠道共生菌赤杆菌Erythrobacter ramosus和脆弱拟杆菌(B. fragilis)通过激活树突状细胞以及在奥沙利铂(oxaliplatin, 一种抗肿瘤药)诱导肠道上皮细胞死亡的情况下释放 IL-1和IL-2,促进滤泡辅助T细胞(follicular T helper cells, Tfh)的诱导。在肿瘤小鼠模型中,这加速了三级淋巴结构中B细胞的成熟,增强了对肿瘤细胞的抗体依赖细胞毒性以及免疫检查抑制剂的效力。然而在复杂情况下精确的微生物-免疫细胞互作的模型还未完全确定。类似的在小鼠模型中,通过表观调控CTL效应分子表达,膳食来源和微生物调控的短链脂肪酸(SCFA)丁酸盐和戊酸盐能够增强靶向表达ROR1肿瘤的CAR-T细胞(chimeric antigen receptor – T cells)对肿瘤的杀伤作用。
外膜囊泡(OMV)是携带微生物物质影响肿瘤免疫的另一种途径。例如,非产毒素的脆弱拟杆菌的OMV能够携带MAMP多糖A(PSA),能够表征局部和全身的抗炎能力。癌症免疫中相似的PSA效应最近被发现,其能通过TLR2信号通路保证CRC细胞增殖并抑制上皮细胞间充质转化。
除了以上描述的远端引发免疫反应的影响,肿瘤微环境中的共生微生物还能够诱导复杂的一系列免疫调节作用,这些作用包括促成主要的免疫抑制环境、通过PRR激活先天性免疫细胞活化促炎的细胞因子产生,从而驱动免疫细胞汇集以及随后的抗原呈递,实现抗肿瘤免疫功能。在某些情况下,相同的调节癌症的微生物可能同时诱导促炎反应和抗炎反应。例如,通过FadA黏附到CRC细胞的具核梭杆菌(F. nucleatum)能够刺激NF-κB、IL-6、IL-8、IL-10、IL-18等炎症因子的释放,增加巨噬细胞、树突状细胞和粒细胞等炎症细胞的渗透,从而造成了一个促炎微环境,促进肿瘤发生。但除此之外,F. nucleatum还能够创造免疫抑制的肿瘤微环境,这可以从CRC中F. nucleatum的丰度与CD3+ T的密度呈负相关反映出来。通过Fap2和RadD诱导T细胞凋亡,或通过Fap2活化抑制性T细胞受体TIGIT可能有利于免疫抑制。此外,F. nucleatum能够释放蛋氨酰-亮氨酰-苯基丙氨酸(formylmethionyl-leucyl-phenylalanine)能够导致MDSC(髓系抑制性细胞)的聚集,并能够调节抑制CD4+ T辅助细胞功能的免疫应答。有趣的是,人类黑色素瘤细胞通过HLA表达一系列呈递在肿瘤细胞表面的肽段,这些肽段来自于包括F. nucleatum在内的肿瘤内细菌。这些微生物肽段抗原呈递能够引起T细胞识别和免疫反应,这表明在某些情况下,肿瘤内驻留的微生物能够直接影响肿瘤微环境的免疫激活。下面将进一步说明微生物组与癌症免疫治疗的相互作用及其对治疗效果的潜在影响。免疫对肿瘤生长和转移形成的潜在影响范围值得进一步研究。
微生物组辅助癌症诊断
随着人们对微生物组可能影响癌症发病机制的认识不断加深,利用个体化微生物来源的数据,将微生物组读数整合到精确的肿瘤场景中,是一个令人兴奋的研究途径。
最近的研究证明,在血液样本中可以检测到独特的微生物DNA和RNA特征。在超过10,000个筛查患者中,使用严格的筛选标准鉴定得到可以预测癌症类型的微生物血浆特征,这些特征能够将其与对应的健康组织表达谱区分开。除了癌症诊断,破解在癌症不同发展阶段的关键微生物特征可能会给分级治疗、转移监测提供可能性。事实上,最近的研究表明,在乳腺肿瘤和其转移部位中,存在具核梭杆菌(F. nucleatum)及其能够上调的肿瘤细胞结合伴侣Gal-GalNAc。相似的,F. nucleatum特征能够在CRC转移灶中检测到,提示F. nucleatum可能有助于转移灶发展。
分析微生物组可能会给预后评估提供可能性。值得注意的是,肿瘤内独特的微生物多样性和组成能够对PDAC患者短期生存和长期生存进行评估。挖掘微生物组数据也可以用于预测癌症免疫疗法和化疗的不良反应。比如,胃肠道和口腔黏膜炎是化疗中主要的并发症,与微生物失调有关。在口腔黏膜炎中,微生物失调的标志是,链球菌(Streptococcus)和普氏菌(Prevotella)属的细菌减少,促炎的F. nucleatum增加。这些菌群失调的特征可以进一步用来评估黑色素瘤患者接受化疗时出现的口腔黏膜炎。
癌症治疗中的微生物组
微生物组除了具有上述的诊断价值外,微生物组还会对癌症治疗反应产生影响,对此进行探索是癌症-微生物组研究中最令人兴奋和最具转化价值的方向之一,并可能优化数据驱动的癌症治疗的决策过程。
免疫治疗
基于免疫的抗癌治疗是指采取一系列旨在增强患者免疫系统或利用免疫成分的方法进行癌症治疗。这种治疗的主要方法是干扰T细胞活化的负调控因子,即免疫检查点,免疫检查点通常会被肿瘤“劫持”,诱导免疫豁免的肿瘤微环境。针对细胞程序性死亡蛋白1(programmed cell death protein 1, PD-1)的抗体或PD-1的配体PD-L1,以及细胞毒性T淋巴细胞相关蛋白4(cytotoxic T lymphocyte-associated protein 4, CTLA-4)的抗体,都是免疫检查点抑制剂(ICI),它们能够阻断T细胞和肿瘤细胞或间质细胞表面抑制性同源配体的互作,从而引发抗肿瘤的免疫反应。在极少数患者中,这种干预能达到完全缓解的效果;甚至在转移性癌症中延长寿命,转移性癌症包括转移性黑色素瘤、非小细胞肺癌、霍奇金淋巴瘤和肾细胞癌等。在2015年,两个小鼠研究显示,包括双歧杆菌属(Bifidobacterium)在内的一些肠道共生微生物能够增强PD-L1ICI抗癌的效果,而多形拟杆菌(Bacteroides thetaiotaomicron)和脆弱拟杆菌(B. fragilis)与增强CTLA-4抑制剂的效果有关。此外,靶向PD-1/L1疗法的抗癌效果与包括Akkermansia、粪杆菌属(Faecalibacterium)、梭菌目(Clostridiales)、双歧杆菌属(Bifidobacterium)在内的多种细菌有关。在无菌小鼠中,合理选择IFNγ诱导的细菌菌株定植后,ICI的效果和抗癌的T细胞反应会显著增强。这些效果部分归因于短链脂肪酸(SCFA)如丁酸和丙酸等微生物代谢物。但是这些影响在一些情况下仍然互相矛盾。例如,粪便SCFA高水平与较长的无进展生存期或增强的抗肿瘤反应有关,但体循环SCFA高水平与较差的治疗反应有关。丁酸盐可能限制树突状细胞诱导肿瘤特异性T细胞和记忆T细胞的能力,从而抑制抗CTLA-4的ICi的疗效。
其他微生物的代谢产物也会影响ICI。比如,长双歧杆菌(Bifidobacterium pseudolongum)产生的肌苷能够通过活化T细胞上的A2A受体增强ICI的作用。在癌症免疫治疗中微生物-宿主相互作用还有其他途径,如Akkermansia muciniphila或拟杆菌通过诱导Th1和CD8+ T细胞的抗肿瘤免疫应答,以IL-12依赖的方式直接刺激淋巴结中的树突状细胞,增强ICI的抗肿瘤疗效。可以根据这些微生物对癌症免疫治疗的影响开发疗法,如使用抗生素、移植微生物或补充代谢物等。
化疗
共生微生物能够调节化疗效果。例如,大肠杆菌可能通过诱导肿瘤耐药和激活细胞毒性调节两种抗癌药物吉西他滨(gemcitabine)和CB1954的疗效。在PDAC患者中吉西他滨可以被细菌代谢,这一效应与肿瘤内LPS的丰度有关,并可以通过抗生素治疗避免药物被代谢。体外实验表明还有十多种其他抗癌药物可能被细菌调节。相反,在小鼠实验中,无菌小鼠或用广谱抗生素处理后的小鼠中奥沙利铂(oxaliplatin)和环磷酰胺(cyclophosphamide)抑制癌症生长的的效果较差。使用抗生素减少共生微生物后,肿瘤浸润髓系细胞对肿瘤免疫治疗的反应较差,可能是使用CpG岛寡核苷酸时肿瘤坏死因子(TNF)生成较少,也可能是奥沙利铂治疗时活性氧减少、细胞毒性不足。此外,化疗相关的肠道屏障损伤会使得肠道共生微生物易位到次级淋巴器官,在小鼠模型中这些微生物能够激活全身Th17型肿瘤特异性抗原CTL(细胞毒性T细胞)的反应。抗生素治疗可以防止肠道共生微生物的易位以及相关的T细胞极化,从而减弱化疗的杀瘤活性。
微生物对化疗效果有影响,化疗和相关的黏膜损伤也会影响肠道微生物的组成。甚至在NGS技术广泛使用之前,基于培养的方法也能为如氟尿嘧啶(5-fluorouracil, 5-FU)等化疗药物能够在实验动物中以增加革兰氏阴性厌氧菌的形式调节口腔和粪便微生物组提供证据。这些发现后来被16S rRNA技术进行了扩展,揭示了真杆菌属(Eubacterium)和瘤胃球菌属(Ruminococcus)的减少。伊立替康(Irinotecan)治疗与特定的肠道微生物菌群失调以及β-葡萄糖醛酸酶表达增加有关。接受骨髓清除治疗的非霍奇金淋巴瘤患者肠杆菌科(Enterobacteriaceae)和肠球菌科(Enterococcaceae)细菌增加,瘤胃菌科(Ruminococcaceae)、毛螺菌科(Lachnospiraceae)和双歧杆菌属减少。异基因造血细胞移植( allogeneic hematopoietic cell transplantation, allo-HCT)和免疫细胞重建与肠道共生微生物粪杆菌属(Faecalibacterium)、瘤胃球菌属(Ruminococcus)、Akkermansia的增加有关。这些化疗引起的微生物改变在肿瘤发生、治疗反应和化疗诱导的不良反应中的影响值得进一步研究。
放疗
放疗与菌群失调的关系有其他人进行了综述。放疗后,肠道微生物失调的特征是,双歧杆菌属(Bifidobacterium)、粪杆菌属(Faecalibacterium)和梭菌属(Clostridium)丰度减少,拟杆菌属(Bacteroides)和肠球菌属(Enterococcus)丰度增加。接受盆腔放疗的患者肠道梭杆菌门(Fusobacteria)增加3%,由于这个门中的一些细菌具有促进肿瘤的能力,可能会带来严重的后果。除去这些关联外,有小鼠研究证实,口服万古霉素清除肠道共生微生物中的革兰氏阳性菌与黑色素瘤、肺癌、宫颈癌放疗疗效的增强有关,这种结果可能是IFNγ和CD8 T细胞依赖机制介导的。相反,在小鼠乳腺癌放疗模型中,抗生素清除包括梭菌目(Clostridiales)等共生微生物,会导致一种肠道酵母增加,促进巨噬细胞介导的促瘤反应。肠道微生物还能通过肠道屏障受损导致的易位调节放疗毒性,即辐射诱导的上皮炎症损伤引起的放射性肠炎,并能进一步引发不可控的肠道免疫反应和组织损伤。同样的,在接受各种局部放疗的患者中,明显的微生物组改变与严重腹泻有关。
调节微生物组改善癌症治疗
微生物组被许多人认为是“第二基因组”,相对人类 基因组其独特之处在于可以调节。合理改变微生物组的干预可能会发展为癌症疗法或癌症并发症的疗法。在接下来的部分中,将举例说明这种实验方法和其在癌症治疗中的潜在用途,并提供了目前正在进行的利用微生物组干预促进癌症治疗的临床研究概述。
微生物移植
通过粪便微生物移植(fecal microbiome transplantation, FMT)将与疾病相关的微生物替换为与健康相关的微生物,在治疗复发性Clostridioides difficile感染和溃疡性结肠炎中非常有效。最近阴道微生物移植在治疗难治性的细菌性阴道病(bacterial vaginosis, BV)方面有了初步进展。将癌症患者的粪便样本转移到经ICI治疗的、无菌或抗生素处理的肿瘤小鼠中,发现特定的微生物组成能够提高免疫疗法的效果。同样的,将对ICI有良好反应的患者的粪便菌群移植到无菌的肿瘤小鼠中,小鼠会对ICI产生反应,但如果将对ICI无反应患者的菌群移植给小鼠,小鼠也会对ICI无反应。最近,两个人体临床试验表明将对ICI有反应的黑色素瘤患者的粪便菌群移植给ICI耐药的黑色素瘤患者,能够逆转一些患者ICI耐药的情况。这些临床实验的效果部分依赖于供体的选择,以及供体微生物是否能成功移植到受体的消化道。FMT尽管已经有了这些初步结果,但在临床方面、监管方面以及科学性方面还有待商榷,比如有效受体和供体的选择、肠道准备以及移植过程等都需要在FMT流程化之前制定标准。此外。驱动FMT发挥临床作用的因素,包括细菌、噬菌体和微生物代谢物等仍旧难以捉摸,值得进一步的大规模、前瞻性临床试验。
益生菌
非处方益生菌制剂被公众广泛使用,但需要有额外的、严格的非行业资助的研究去评估益生菌的有效性、定植能力和可能的副作用。益生菌制剂对癌症的发病有影响,但在不同病例中会出现相互矛盾的结果。例如,罗伊氏乳杆菌(Lactobacillus reuteri)与幽门螺杆菌(H. pylori)具有竞争关系,补充罗伊氏乳杆菌会显著降低幽门螺杆菌的数量,这表明高度靶向的细菌补充剂可能起到预防癌症的作用。在氧化偶氮甲烷/右旋糖酐硫酸钠(azoxymethane/dextran sulfate sodium, AOM/DSS)处理的CRC小鼠模型,以及1,2-二甲肼二盐酸化物(1,2-dimethylhydrazine dihydrochloride, DMH)处理的CRC大鼠模型中,乳酸杆菌(Lactobacilli)和双歧杆菌(Bifidobacteria)能够通过增加短链脂肪酸(SCFA)的生成,诱导细胞凋亡、抑制癌症细胞增殖,从而减少癌症的发生和进展、减小肿瘤体积。此外,一些益生菌能够改善化疗、放疗的副作用。相反的是,一些研究表明给癌症患者服用益生菌补充剂收效甚微或没有作用,甚至会引发一些不良反应。例如,一项临床试验显示,急性髓系白血病患者接受化疗后,补充罗伊氏乳杆菌和鲍氏酵母菌(Saccharomyces boulardii),也没有显著改变免疫抑制。此外,这种益生菌补充剂实际上增加了患者得的全身感染率。同样,补充乳酸菌(Lactobacillus)几乎不能控制子宫内膜腺癌患者接受放疗后出现的腹泻,补充Lactococcus brevis也不能改善头颈癌患者的口腔黏膜炎。值得注意的是,益生菌的使用可能会对原有微生物造成严重影响,包括抑制抗生素治疗后的微生物重建、增强细菌的抗生素耐药等;而且在一些情况下,全身感染或局部感染在免疫抑制的癌症环境中可能是危险的。
这些不同的、有时矛盾的结果可能是由于原有微生物对外源微生物的定植抗性具有个体化差异。开发新的”精准益生菌”制剂优化肠道定植能力,可重现对癌症的反应,同时保证病人的安全性,是一个令人兴奋的活跃的研究领域。这些研究整合了表型筛选方法,包括对微生物分泌的小分子对肿瘤和免疫细胞的影响的解释,具有新生物活性的“精准益生菌”的芯片预测,以及微生物工程。这种精准益生菌制剂可以根据微生物组成和临床特征针对不同个体使用,并可以作为癌症治疗的辅助剂。
微生物靶向肿瘤
除了口服微生物群落或分离出的益生菌,肿瘤微生物作为潜在的肿瘤导向载体进行系统给药的设想已经持续了几十年。利用沙门氏菌(Salmonella)的减毒突变株作为迁移-吸引成分进行了临床Ⅰ期试验,但缺乏肿瘤特异性。在小鼠模型中微生物经过进一步修饰,通过激活NLRP3炎症小体以及诱导IL-1B、IL-18和TNFα,能够提高肿瘤特异性和抗肿瘤活性。最近,一种鸟苷四磷酸(guanosine tetraphosphate, ppGpp)减毒沙门氏菌可以在小鼠中激活肿瘤微环境中的先天性免疫和促炎通路,导致整体的肿瘤血管系统被破坏。
此外,肠道微生物中可以被移植的共生菌可以作为合成生物学底盘微生物。细菌已经进化出广泛的具有潜在用途的生理特性和代谢特性,这些特性可以被用来调节肿瘤微环境。例如,在小鼠实验中,基因工程改造的鼠伤寒沙门氏菌(Salmonella typhimurium)表达溶血素E(hemolysin E),且群体感应通路被改变,能够有效靶向肿瘤位点并使细胞裂解,从而降低肿瘤活性,并提高生存率。益生菌大肠杆菌Nissle 1917经过修饰能够产生纳米级抗体,在肿瘤内靶向PD-L1和CTLA-4,具有很好的减小肿瘤的能力。一种非致病性的大肠杆菌菌株被基因工程改造表达天青蛋白(azurin),在小鼠模型中这种细菌蛋白能诱导肿瘤细胞凋亡,防止肿瘤转移,或者在肿瘤微环境中释放抗CD47拮抗物的纳米级抗体,激活肿瘤浸润T细胞使肿瘤细胞裂解。婴儿双歧杆菌(Bifidobacterium infantis)重编程胞嘧啶脱氨酶,无毒性的5-氟胞嘧啶转化胃具有细胞毒性的化合物5-FU。在小鼠模型中,给予药物前体以及表达脱氨酶的婴儿双歧杆菌能够抑制黑色素瘤的生长。为了避免细菌诱导的和肿瘤细胞裂解诱导的全身免疫反应,以及由此引发的细胞因子释放综合症,会在工程菌中设置切断开关(kill switch),控制细菌在宿主中的复制以及活性的持续时间,同时限制潜在的毒性。在保证患者安全的同时,需要严格的临床试验评估在癌症情况下,移植微生物的有益效果。
日常饮食
改变饮食结构是将微生物群落重塑成健康的组成是一种吸引人的方式,这些微生物主要是但并不局限于肠道微生物,同时也能提高癌症治疗的有效性。“西方的”饮食富含脂肪、精制碳水和果糖,可能导致CRC、前列腺癌和乳腺癌的发生。在小鼠中,将其饮食转变为高脂饮食后,会增加厚壁菌门(Firmicutes)和变形菌门(Proteobacteria)的丰度,同时降低拟杆菌(Bacteroidetes)的丰度,并激活NF-κB炎症途径,与癌症的发病有关。与之相反的是,高纤维饮食在小鼠模型中会诱导细菌产生乙酸、丙酸、丁酸等短链脂肪酸(SCFA),这些SCFA都与抗炎和促进凋亡的特性。给腺癌小鼠口服一种名为菊粉(inulin)的多糖膳食纤维,能够显著增强抗PD-1的疗效,增加肠道中的Akkermansia以及与增加免疫检查点抑制剂(ICI)有关的SCFA。菊粉能够改变TME中的免疫环境,PD-1+ CD8+ T细胞增加,调节性T细胞减少。
然而这些研究的大多数结果是在实验动物中进行的,而忽略了现实生活中人生理状态和疾病相关反应的个体差异。近年来,个性化营养项目(Personalized Nutrition Project)和PREDICT1研究报告称,来自两个大型人群队列的微生物组和宿主特征可以被用来开发基于机器学习的餐后血糖血脂反应的个性化预测。事实上,个性化预测的饮食调整可以短期或长期减少糖尿病前期患者饭后血糖峰值的代谢后果。同样利用癌症患者的饮食反应,可以定制适合于该个体及其微生物的饮食,优化治疗反应。这些饮食调整方法是正在进行的、令人兴奋的研究途径。
后生元疗法
未确定的微生物生物活性化合物的混合物被认为在某些癌症中具有生物活性,比如胞外多糖(exopolysaccharide, EPS)制剂和乳酸菌(Lactobacillus)的无细胞上清(cell-free supernatants, CFS)。在体外,EPS诱导肝脏和消化道肿瘤细胞系增殖减少,来自乳酸菌、双歧杆菌(Bifidobacterium)和粪杆菌(Faecalibacterium)的CFS诱导细胞凋亡、减少肿瘤细胞增殖,激活抗炎信号通路。同样的,细菌和酵母细胞壁中的一组复杂的葡萄糖聚合物葡聚糖(beta-glucans)能够通过C3b补体包被肿瘤细胞增强白细胞对肿瘤细胞的杀伤作用。
后生元疗法即指通过明确定义微生物生物活性化合物和它们的补剂改进这些方法进行癌症治疗。例如,选定拟杆菌目(Bacteroidales)、真杆菌属(Eubacterium)、粪杆菌属(Faecalibacterium)定植的无菌小鼠肠道和血清中含有丰富的甲羟戊酸(mevalonate)和二甲基甘氨酸(dimethylglycine),与增强抗肿瘤免疫和ICI疗效有关。吲哚是细菌色氨酸代谢的主要产物,且是芳基烃受体的配体,在黏膜稳态中起着重要作用。的确,卵巢癌患者血清中的3-吲哚丙酸的水平下降。体外实验中吲哚具有乳腺癌细胞株的抑制活性,在小鼠乳腺癌模型中吲哚补剂可减少癌症转移。
另一种后生元疗法是利用外膜囊泡(OMV)作为肿瘤调节物质的运输载体。经修饰的大肠杆菌OMV在结直肠癌小鼠中可作为免疫疗法的药物,其通过在肿瘤中积累并产生IFNγ增强肿瘤微环境中的抗肿瘤反应。一种抗原修饰的OMV疫苗能够引发特异性的抗肿瘤免疫反应,避免肺黑色素瘤转移、抑制皮下CRC生长。增加对饮食、宿主、调节癌症的微生物小分子和OMV的了解,可能是未来十年癌症-微生物研究中重要的方面。
抗生素治疗
在癌症中利用抗生素清除微生物的方法仅应用于通过清除幽门螺杆菌预方胃癌和MALT淋巴瘤。因为考虑到幽门螺杆菌是一种共生菌,且在多数情况下与癌症发展无关,因此即使在这种特殊情况下根除幽门螺杆菌仍有争议。在实验室小鼠皮下注射MC-26癌细胞构建的模型中,一些实验性抗生素能够靶向代谢吉西他滨的细菌,从而保证吉西他滨的疗效。最近的一项Ⅲ期临床试验在评估这种方法应用于PDAC治疗的可行性。然而广谱抗生素治疗在肿瘤进展中存在有害影响。对于使用同种异体造血细胞移植治疗的恶性血液病患者以及使用ICI的晚期黑色素瘤患者,抗生素会造成双歧杆菌或Akkermansia减少,与其相关的SCFA等微生物代谢物也会减少,共同作用降低治疗效果。同样的,在使用ICI治疗前30天使用广谱抗生素的患者,相比较晚时间点使用抗生素或不使用抗生素的患者,总生存率显著降低。由于抗生素是癌症患者对抗感染、保证生存的治疗手段,目前正在研究将抗生素对内源性微生物影响降到最低的策略。一项Ⅲ期临床试验在急性髓系白血病患者或骨髓增生异常综合征患者中评估了靶向结肠的抗生素吸附剂。总的来说,由于抗生素对患者体内微生物的影响无差别,使用抗生素清除促进癌症的病原体仍具有局限性,可能会导致不良反应、耐药菌株的出现以及一些对病程和治疗效果不可预见的影响。靶向清除促进癌症的共生微生物,同时对微生物造成影响尽可能小的新方法是一个活跃的研究领域。
噬菌体疗法
噬菌体,即能够感染细菌的病毒,在自然界中无处不在,并可能通过“噬菌体爆发”(phage bloom)对肠道微生物的组成造成影响。噬菌体作为靶向的抗菌治疗手段一直在被改进。全身给药单一噬菌体或噬菌体鸡尾酒疗法在小鼠和人类的细菌感染中有初步疗效。噬菌体疗法的挑战包括细菌的噬菌体耐性、潜在的免疫原性、补体激活以及肝脏和脾脏吞噬作用介导的噬菌体快速清除。口服噬菌体给药可能会避免全身免疫,同时靶向疾病相关的肠道共生微生物,而对其他微生物没有太多影响。实际上,使用去除细菌的病毒样颗粒(90%是噬菌体)的粪便微生物移植能够改变小鼠高脂饮食诱导的菌群失调,且被证明在人体中可行,尽管FMT受体的个体定植差异是显著的。通过多种受体靶向疾病相关病原体的更明确的噬菌体鸡尾酒疗法可能增强病原体靶向的特异性,并能防止噬菌体抗性的出现。然而这种方法面临许多挑战。尽管缺乏宿主噬菌体受体,先天性免疫系统可能感知噬菌体DNA,从而在某情况下引起免疫反应。此外,胃肠道噬菌体的输送可能会由于胃酸、肠道黏膜层以及相关生物膜结构等生物物理环境而变得复杂,可能会导致噬菌体的失活或限制噬菌体与细菌靶点的结合。
在癌症中,使用噬菌体制剂可能会靶向清除促进癌症的共生微生物,同时对周围微生物的影响很小。初步研究表明,在体外模型中,种群特异性裂解的噬菌体能够清除癌症相关的、共生的幽门螺杆菌。此外,运载药物的治疗型噬菌体能够直接靶向驻留在癌症中的细菌,在肿瘤微环境中释放药物。例如经过设计的具核梭杆菌(F. nucleatum)特异性噬菌体能够携带伊立替康(irinotecan)纳米粒子,当噬菌体迁移到F. nucleatum聚集的CRC位点时,就能在肿瘤微环境中释放纳米粒子。
微生物组和癌症研究中的挑战
随着对微生物组及其对健康和疾病影响的进一步探究,大量计算数据和实验数据的积累有助于我们理解包括癌症等的疾病过程,同时利用这些知识可以开发新的诊断方式和治疗方式。然而,这种多学科研究面临重大的技术挑战和概念障碍,因此为了获得精确可行地理解宿主-微生物的相互作用对健康和疾病的影响,应该认识到并尝试解决这些障碍。
样本选择
大多数微生物组研究利用粪便样本或口腔样本作为胃肠道微生物的可获得代表。但是这样生态位相关性会受到粪便和胃肠道中微生物组成的内在差异的影响。依据粪便样本计算肠道微生物群落结构,或直接通过微创方法取样肠道微生物,可能会优化微生物群落评估的准确性和可重复性。更加困难的是评估人类肿瘤微生物。肿瘤内微生物信号虽然难以获得但具有临床价值。例如,胰腺癌患者进行肿瘤手术时的肿瘤微生物多样性可能可以预测生存率。获取这些样本并将其作为临床决策库的一部分是具有挑战性的,但在未来一些年会成为令人兴奋的研究领域。
数据和资源的可用性
二代测序技术(NGS)、代谢组学和蛋白质组学的进展使得用于识别癌症进展、抗癌治疗效果相关的微生物组成和功能特征的高通量数据的产生成为可能。在这一方面的主要挑战是数据和资源的可用性,由于普遍的科学可重复性危机,可用性是优化微生物组研究的关键要求。另一个挑战是数据采集、数据分析方法和技术(如DNA提取技术、16S rRNA基因的PCR)的不统一,限制了比较、整合和探索来自不同研究、不同地理区域的数据集,同时也限制了对结果的归纳。不过令人鼓舞的是,越来越多以微生物组为主的论文要求测序数据和详细方法可用通过欧洲核酸库(European Nucleotide Archive)或NIH序列库(Sequence Read Archive of NIH)公开获取。然而这些数据往往缺少足够详细的相应的元数据(metadata),部分是由于地方机构的审查委员会会加以限制,尽管患者身份已经去标识化。同样的,样本大小和统计计算方法的共享没有充分实施。目前网页应用可以达到这些目的(如https:// fedematt.shinyapps.io/shinyMB/)。未来十年微生物组研究需要更加统一,做好数据共享以及生物信息工具的共享,从而实现更统一的去噪、宿主读序去除、分类以及基因数据库比对。
个体间微生物多样性
除了以上所述的技术差异会对微生物组结果造成影响外,个体微生物组成的独特性导致的个体间和个体内微生物差异,是区分真实信号和噪声、归纳结果面临的巨大挑战。例如,有三个近期发表的具有里程碑意义的研究将肠道微生物组作为ICI疗效的预测因子,揭示了不同类群与免疫治疗结果的相关性。微生物组数据极具可变性,经常受到质疑,但是这可能是真实的生物可变性而不是方法论的缺陷。包括地理环境、年龄、性别、生活习惯、遗传因素和潜在疾病等在内的多种宿主因素和环境因素影响着个体间微生物组的差异。重要的是,个体间微生物组差异并不一定是不可分析的,反而可以用来识别在遗传风险相似但临床表现不同的情况下,个体特异性的、疾病背景特异性的微生物组成。这种个性化的微生物组标签可以说是一种大数据指纹,可能有助于利用机器学习去预测个性化的生理、癌症相关的特征和反应。为了实现这种个性化分析,可能需要新的计算工具来准确获取个体特异、疾病特异的微生物组随时间变化的模式,以及这期间微生物组对临床指标的影响。
相关性和因果关系
癌症-微生物研究领域最大的挑战之一是超越关联性和相关性的分析,明确因果关系和机制。在证明微生物组因果关系中广泛使用的方法是将整个微生物组、确定的菌群或单种微生物移植到无菌小鼠中,使得研究人员能模拟微生物对人类癌症和治疗的影响。在一项研究中,给予小鼠致癌剂氧化偶氮甲烷(azoxymethane, AOM),移植CRC患者的粪便微生物的小鼠比移植健康对照组粪便微生物的小鼠要有更高比例的Ki67阳性的增殖细胞以及炎症标志物。此外,与对照组相比,将CRC患者粪便微生物移植到常规小鼠中,会夹中息肉和高度肠道发育不良。同样,对从人结肠息肉生物膜上分离出的、包含colibactin合成必需的pks毒力岛的产肠毒素脆弱拟杆菌(B. fragilis)和大肠杆菌(E. coli)进行单独移植,会加速AOM和APCMin/+肿瘤小鼠模型肠道肿瘤的发展。
除了小鼠模型中的体内实验,体外模型也是检测微生物及其分泌的分子对癌症直接或间接影响的重要方法。例如,微流控肠道芯片模型可用于厌氧条件下共培养人类肠道上皮细胞和共生微生物,以研究炎症过程或宿主-病原体的相互作用。小肠或大肠生成的三维器官模型能够解析不同共生微生物及其生物活性产物对癌症发生和进展的影响机制。例如,聚核梭杆菌(F. nucleatum)与人类肿瘤器官共培养可以诱导参与癌症转移途径的基因表达。人类肠道器官与分泌colibactin的大肠杆菌共培养下,发现了人类CRC单碱基替换的变异特征,因此揭示了共生微生物促进癌症的基因毒性。新方法的发展可以将免疫细胞、其他肿瘤微环境细胞以及复杂的微生物群落整合到同一类器官环境中。实际上,在短期的3D培养中,保留免疫细胞的来自病人的肿瘤能够对免疫检查点抑制剂产生反应。
结论
在不到十年的时间里,微生物组领域为人类生理学和疾病研究提供了有价值的见解。人类癌症是最复杂、最具破坏性、最缺乏了解的疾病之一,其与特定微生物的改变以及微生物群落结构整体的改变有关。研究黏膜部位以及肿瘤微环境中共生微生物的因果关系和分子相互作用,能够为揭示癌症发生、发展和治疗反应的差异提供新的线索。除了尝试从相关性的解释走向因果性的解释外,研究微生物组对癌症的影响还面临着样本选择、处理、测序和数据分析相关的艰巨挑战。尽管存在这些挑战,随着微生物对癌症诊断、病人分级和治疗的影响越来越大,微生物对癌症的影响可能会在未来十年的癌症研究中占据中心地位。
关注“药时代”微信公众号,获取更多精彩内容 ↓ ↓ ↓
End

推荐阅读

 点击这里,与~20万同药们喜相逢!
点击这里,与~20万同药们喜相逢!本篇文章来源于微信公众号:药时代
发布者:药时代,转载请首先联系contact@drugtimes.cn获得授权
 为好文打赏 支持药时代 共创新未来!
为好文打赏 支持药时代 共创新未来! 